

- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы
- •Под редакцией ю. Ашоффа
- •В двух томах
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Предисловие редактора перевода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Предисловие
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Список авторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 1. Обзор биологических ритмов ю. Ашофф
- •Спектр ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Четыре «циркаритма»
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •16 Глава 1
- •Ультрадианные и инфрадианные ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Взаимодействие ритмов и их целесообразность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •«Цирка»-осцилляторы в качестве часов
- •Распознавание местного времени: захватывание колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Измерение промежутков времени: гомеостаз периода н угловой скорости колебателя
- •Происхождение и дифференциация функции колебателя как биологических часов Циркадианные программы: от экзогенной упорядоченности к эндогенной организации
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Период колебателя и время генерации
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •«Оппортунистическая» эволюция: дифференциация функций колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Колебатель и программа Одноклеточные системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •32 Глава 2
- •Местонахождение колебателя в многоклеточных системах
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •34 Глава 2
- •Локализация колебателя и проводящие пути
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Взаимное сопряжение нескольких колебателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Колебатель и подневольные ритмы: иерархия захватывания
- •Колебатель и подневольные ритмы: периодическая программа
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Искажение периодической программы внешними световыми циклами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Фотопериодизм: сезонные изменения периодической программы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •48 Глава 2
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Единство в многообразии циркадианных систем
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Свободнотекущие ритмы Зависимость периода от внешних н внутренних факторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Изменчивость периода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захваченные ритмы Многообразие принудителей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Условия захватывания и соотношение фаз
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиг фазы принудителя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Некоторые элементы теории колебаний
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Динамические модели-циркадианного колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •76 Глава 4
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Обзор моделей, описывающих действие непрерывного освещения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •78 Глава 4
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Толкования динамической модели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Явления в популяциях осцилляторов
- •Общие свойства популяций осцилляторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание: общие принципы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Два механизма захватывания — дискретный и непрерывный
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Кривые смещения фазы (ксф) циркадианных колебателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Кинетика сдвига фазы колебателя: поведение предельного цикла
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Дискретное захватывание колебателя внешними световыми циклами Фаза колебателя относительно принудителя в устойчивом захваченном состоянии
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пределы захватывания: воспроизведение периода
- •Умножение периода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание «скелетными» фотопериодами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание «полными» фотопериодами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Стабильность устойчивого захваченного состояния Скелетные фотопериоды
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сезонные изменения длины дня: и форма ксф
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сезонные изменения длины дня: составные колебатели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание отдельного колебателя и захватывание системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •120 Глава 5
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Перспективы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы локомоторной активности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы питания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы спаривания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы репродуктивного поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Поведение, ограниченное «воротами»
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •136 Глава 6
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы ориентационного поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Чувство времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмичность в процессах научения
- •Ритмические изменения реактивности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •142 Глава 6
- •Общая циркадианная организация поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианные колебатели в нервной системе Колебатели in vitro
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Определение локализации путем пересадки
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Выяснение .Локализации путем повреждений
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •164 Глава 7
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Множественные колебатели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианная ритмичность и сенсомоторная интеграция Циркадианная модуляция активности центральной нервной и нейросекреторной систем
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианная модуляция сенсорных сигналов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Фоторецепция и захватывание Локализация фоторецепторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •178 Глава 7
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Нервные механизмы захватывания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Генетика колебателя и наблюдаемых ритмов
- •Мультигенный анализ
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Анализ отдельных генов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •196 Глава 8
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Млекопитающие
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •208 Глава 9
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Научение и память
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Птицы Введение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пресмыкающиеся
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Земноводные
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рыбы Введение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •226 Глава 9
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарности
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 10. Внутренняя временная упорядоченность
- •Внутренняя временная упорядоченность при устойчивом захваченном состоянии
- •Факторы, определяющие форму колебаний
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Карты фаз циркадиаиных систем
- •Наследственные компоненты внутренней временной упорядоченности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пластичность фазы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сила сопряжения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Временная упорядоченность в отсутствие внешней информации о времени Внутренняя согласованность ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиги внутренних фазовых отношений
- •Изменения формы циркадианных колебаний
- •Повышенная пластичность фазы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Анатомические и физиологические предпосылки внутренней временной упорядоченности
- •Абстрактные модели внутренней организации циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Качественные модели циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рецепция сигналов от времязадателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Значение внутренней временной упорядоченности
- •Преимущества периодической организации процессов жизнедеятельности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Последствия нарушений строгой временной упорядоченности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Восприятие захватывающих сигналов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •280 Глава 11 Первичные ритмоведущие структуры
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Эпифиз птиц
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Супрахиазменные ядра млекопитающих
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Затухающие осцилляторы и ведомые ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Возникновение организации
- •Измерение зарождающихся ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Развитие наблюдаемой ритмичности у человека
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Физиология появления ритмичности у крыс
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •300 Глава 12
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Развитие циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Роль внешней среды в онтогенезе ритмов
- •Циклы освещения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Мать как циркадианный времязадатель
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Внутренняя среда
- •Старение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Перспективы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарность
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 13. Ежедневные приспособительные стратегии поведения
- •Введение: давление отбора и суточные ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень сообщества: ночной и дневной образ жизни
- •Роль дня и ночи в эволюционной дивергенции
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Специализация во времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Разделение во времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиги временной ниши
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень вида: распределение активности на протяжении суток
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Добывание и потребление пищи
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ежедневные перемещения и миграции
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Размножение и онтогенез
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень индивидуума: ежедневные повадки
- •Память на время у медоносной пчелы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •338 Глава 13
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Предвосхищение периодического появления пищи
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Стратегия повадок
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические часы и ориентация по небесным светилам
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Солнце как ориентир
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Луна как ориентир
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Звездные ориентиры
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Другие аспекты периодических изменений ориентированной активности
- •Периодические изменения направления
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Периодическое изменение передвижений
- •Благодарности
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Свободнотекущие ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание искусственными принудителями
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Внутренняя десинхронизация и частичное захватывание
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Эксперименты со сдвигом фазы и перелеты в широтном направлении
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Приложения в области медицины
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарность
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рекомендуемая дополнительная литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Зависимость от времени дня
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Круглосуточные исследования
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Влияние сдвига фазы времязадателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Мотивация, обстановка и индивидуальные различия
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Оглавление
- •Глава 1. Обзор биологических ритмов. Ю. Ашофф ... 12
- •Глава 2. Циркадианные системы: общая перспектива.
- •Глава 3. Свободнотекущие и захваченные циркадианные ритмы. Ю. Ашофф 54
- •Глава 4. Математические модели. Т. Павлидис .... 70
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Глава 5. Циркадианные системы: захватывание. К. Пит-
- •Глава 6. Ритмы поведения у беспозвоночных. Дж. Брэйди 12s
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Глава 16. Ритмы работоспособности. П. Колькюхунь ... 389
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Уважаемый читатель!
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Электронное оглавление
- •Глава 1. Обзор биологических ритмов 12
- •Глава 2. Циркадианные системы: общая перспектива 22
- •Глава 3. Свободнотекущие и захваченные циркадианные ритмы 54
- •Глава 4. Математические модели 70
- •Глава 5. Циркадианные системы: захватывание 87
- •Глава 6. Ритмы поведения у беспозвоночных 125
- •Глава 7. Нервный и эндокринный контроль циркадианной ритмичности у беспозвоночных 152
- •Глава 8. Генетика и развитие циркадианных ритмов беспозвоночных 188
- •Глава 9. Ритмы поведения позвоночных 200
- •Глава 10. Внутренняя временная упорядоченность 240
- •Глава 11. Нервный и эндокринный контроль циркадианных ритмов у позвоночных 275
- •Глава 12. Онтогенез циркадианных ритмов 292
- •Глава 13. Ежедневные приспособительные стратегии поведения 315
- •Глава 14. Ориентация в пространстве с помощью часов 348
- •Глава 15. Циркадианная система человека 362
- •Глава 16. Ритмы работоспособности 389
Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
Ритмы поведения у беспозвоночных 141
|
|
|
Рис. 3. Сравнение изменений поведенческого и рецепторного порогов на протяжении циркадианного цикла у двух насекомых при СТ 12 : 12, как указано на оси абсцисс. А. Изменение реакции самцов бабочки Trlchoplusia ni на синтетический половой феромон самки. Черные кружочки — доза, необходимая для появления электрической реакции антенн (средние величины для 7 — 10 самцов). Светлые кружочки — доза, вызывающая поведенческую реакцию у 50% самцов. Последние две точки обеих кривых представлены иа графике дважды. (По [81].) Б. Изменение реакции самцов мясной мухи Protophormia terraenovae к стимуляции лапок сахарозой. Черные квадратики и левая ордината — доля мух (n=83), вытягивающих хоботок при воздействии 1,0 M сахарозы. Светлые квадратики и правая ордината — концентрация сахарозы, вызывающая реакцию у 50% мух (n=181) (порог чувствительности к сахарозе; шкала логарифмическая!). Кружочки — число спайков, возникающих в одном тарзальном хеморецепторном волоске в первые полсекунды при стимуляции 0,3 M сахарозой (17 мух; каждый раз регистрировались ответы двух волосков). Вертикальными отрезками указана стандартная ошибка. СТ 12:12, на оси абсцисс представлена только часть темной фазы. (По [50].) |
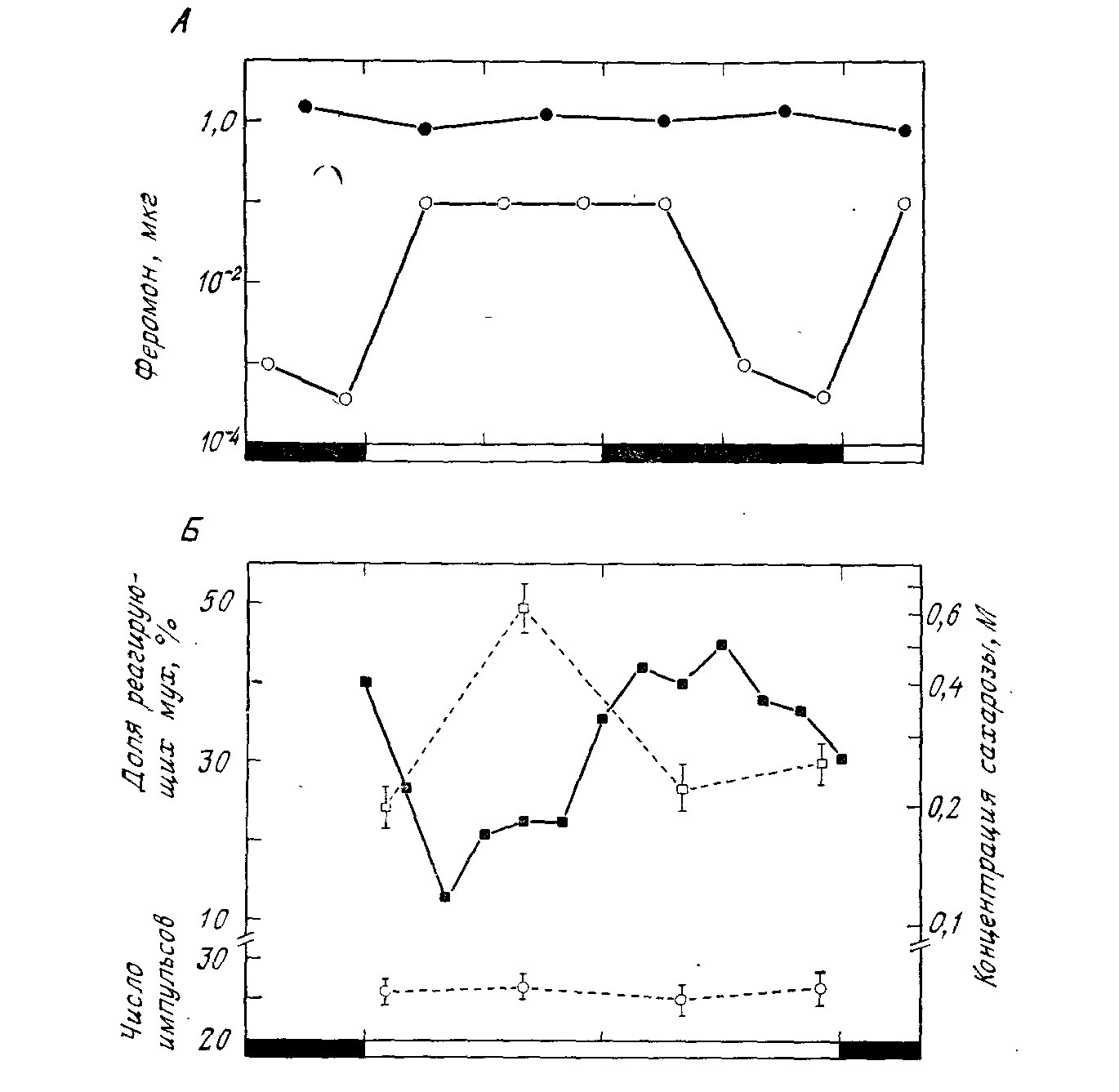
Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
142 Глава 6
звуку полета самок (см. выше). С этой точки зрения были изучены еще три поведенческих ритма, но ни в одном случае не было обнаружено аналогичных ритмов чувствительности рецепторов. Первый из этих случаев — ритм чувствительности бабочек-самцор к половому феромону самки. У Trichoplusia ni происходят четкие циркадианные изменения в пороге поведенческой реакции, но нет соответствующих изменений в электроантеннограмме (рис. 3,А); так же обстоит дело у самцов сатурний [89]. Второй случай — рефлекс вытягивания хоботка на раздражение лапок сахаром у мясных мух. И здесь обнаружен ясный циркадианный ритм в поведенческой реактивности (в этом случае определяли либо пороговую концентрацию сахара, либо долю реагирующих мух), но не было никаких циркадианных изменений в афферентных сигналах от хеморецепторов лапок (рис. 3, Б); этот ритм, кроме того, проходит не менее двух циклов при постоянном освещении [50]. Третий, менее убедительный, случай — это реакция мухи цеце на медленно движущиеся зрительные стимулы. Здесь снова имеется четкий циркадианный ритм поведенческой реактивности (рис. 4, B), но косвенные психофизические данные позволяют думать, что он не обусловлен изменениями в чувствительности глаз [21].
Таким образом, в этих трех случаях есть веские основания полагать, что ритмы поведенческой реактивности связаны не с периферическими ритмами чувствительности рецепторов, а с ритмами в процессах центральной интеграции соответствующих афферентных сигналов.
Общая циркадианная организация поведения
Для циркадианных поведенческих ритмов, по-видимому, характерно то, что у каждого данного вида многие формы поведения совпадают по фазе. В этом отношении наиболее подробно изучена муха цеце (рис. 4), но совпадающие по фазе циркадианные ритмы существуют также у дождевых червей (способность к научению, локомоция, отрицательный фототаксис [2, 14]), гребляков (локомоция и фототаксис [фотокинез?] 88]), водомерок (локомоция, геотаксический угол [15]), комаров (полет, спаривание, яйцекладка [56, 49]), бабочек (чувствительность к феромону, полет [8]), а также у позвоночных — птиц [77] и человека [6].
Совпадение некоторых из этих ритмов, вероятно, обусловлено прямым или косвенным влиянием ритма локомоции (см. выше), но во многих случаях соответствующие формы поведения связаны с различными сенсорными системами, как, например, у дождевых червей, водомерок и мух цеце. Поэтому кажется вероятным, что параллелизм в поведенческой циркади-