

- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы
- •Под редакцией ю. Ашоффа
- •В двух томах
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Предисловие редактора перевода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Предисловие
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Список авторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 1. Обзор биологических ритмов ю. Ашофф
- •Спектр ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Четыре «циркаритма»
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •16 Глава 1
- •Ультрадианные и инфрадианные ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Взаимодействие ритмов и их целесообразность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •«Цирка»-осцилляторы в качестве часов
- •Распознавание местного времени: захватывание колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Измерение промежутков времени: гомеостаз периода н угловой скорости колебателя
- •Происхождение и дифференциация функции колебателя как биологических часов Циркадианные программы: от экзогенной упорядоченности к эндогенной организации
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Период колебателя и время генерации
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •«Оппортунистическая» эволюция: дифференциация функций колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Колебатель и программа Одноклеточные системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •32 Глава 2
- •Местонахождение колебателя в многоклеточных системах
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •34 Глава 2
- •Локализация колебателя и проводящие пути
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Взаимное сопряжение нескольких колебателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Колебатель и подневольные ритмы: иерархия захватывания
- •Колебатель и подневольные ритмы: периодическая программа
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Искажение периодической программы внешними световыми циклами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Фотопериодизм: сезонные изменения периодической программы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •48 Глава 2
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Единство в многообразии циркадианных систем
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Свободнотекущие ритмы Зависимость периода от внешних н внутренних факторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Изменчивость периода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захваченные ритмы Многообразие принудителей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Условия захватывания и соотношение фаз
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиг фазы принудителя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Некоторые элементы теории колебаний
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Динамические модели-циркадианного колебателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •76 Глава 4
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Обзор моделей, описывающих действие непрерывного освещения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •78 Глава 4
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Толкования динамической модели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Явления в популяциях осцилляторов
- •Общие свойства популяций осцилляторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание: общие принципы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Два механизма захватывания — дискретный и непрерывный
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Кривые смещения фазы (ксф) циркадианных колебателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Кинетика сдвига фазы колебателя: поведение предельного цикла
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Дискретное захватывание колебателя внешними световыми циклами Фаза колебателя относительно принудителя в устойчивом захваченном состоянии
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пределы захватывания: воспроизведение периода
- •Умножение периода
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание «скелетными» фотопериодами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание «полными» фотопериодами
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Стабильность устойчивого захваченного состояния Скелетные фотопериоды
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сезонные изменения длины дня: и форма ксф
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сезонные изменения длины дня: составные колебатели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание отдельного колебателя и захватывание системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •120 Глава 5
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Перспективы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы локомоторной активности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы питания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы спаривания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы репродуктивного поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Поведение, ограниченное «воротами»
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •136 Глава 6
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмы ориентационного поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Чувство времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ритмичность в процессах научения
- •Ритмические изменения реактивности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •142 Глава 6
- •Общая циркадианная организация поведения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианные колебатели в нервной системе Колебатели in vitro
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Определение локализации путем пересадки
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Выяснение .Локализации путем повреждений
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •164 Глава 7
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Множественные колебатели
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианная ритмичность и сенсомоторная интеграция Циркадианная модуляция активности центральной нервной и нейросекреторной систем
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Циркадианная модуляция сенсорных сигналов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Фоторецепция и захватывание Локализация фоторецепторов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •178 Глава 7
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Нервные механизмы захватывания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Генетика колебателя и наблюдаемых ритмов
- •Мультигенный анализ
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Анализ отдельных генов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •196 Глава 8
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Млекопитающие
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •208 Глава 9
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Научение и память
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Птицы Введение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пресмыкающиеся
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Земноводные
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рыбы Введение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Активность
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •226 Глава 9
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарности
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 10. Внутренняя временная упорядоченность
- •Внутренняя временная упорядоченность при устойчивом захваченном состоянии
- •Факторы, определяющие форму колебаний
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Карты фаз циркадиаиных систем
- •Наследственные компоненты внутренней временной упорядоченности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Пластичность фазы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сила сопряжения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Временная упорядоченность в отсутствие внешней информации о времени Внутренняя согласованность ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиги внутренних фазовых отношений
- •Изменения формы циркадианных колебаний
- •Повышенная пластичность фазы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Анатомические и физиологические предпосылки внутренней временной упорядоченности
- •Абстрактные модели внутренней организации циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Качественные модели циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рецепция сигналов от времязадателей
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Значение внутренней временной упорядоченности
- •Преимущества периодической организации процессов жизнедеятельности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Последствия нарушений строгой временной упорядоченности
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Восприятие захватывающих сигналов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •280 Глава 11 Первичные ритмоведущие структуры
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Эпифиз птиц
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Супрахиазменные ядра млекопитающих
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Затухающие осцилляторы и ведомые ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Возникновение организации
- •Измерение зарождающихся ритмов
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Развитие наблюдаемой ритмичности у человека
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Физиология появления ритмичности у крыс
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •300 Глава 12
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Развитие циркадианной системы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Роль внешней среды в онтогенезе ритмов
- •Циклы освещения
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Мать как циркадианный времязадатель
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Внутренняя среда
- •Старение
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Перспективы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарность
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Глава 13. Ежедневные приспособительные стратегии поведения
- •Введение: давление отбора и суточные ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень сообщества: ночной и дневной образ жизни
- •Роль дня и ночи в эволюционной дивергенции
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Специализация во времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Разделение во времени
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Сдвиги временной ниши
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень вида: распределение активности на протяжении суток
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Добывание и потребление пищи
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Ежедневные перемещения и миграции
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Размножение и онтогенез
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Уровень индивидуума: ежедневные повадки
- •Память на время у медоносной пчелы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •338 Глава 13
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Предвосхищение периодического появления пищи
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Стратегия повадок
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические часы и ориентация по небесным светилам
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Солнце как ориентир
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Луна как ориентир
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Звездные ориентиры
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Другие аспекты периодических изменений ориентированной активности
- •Периодические изменения направления
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Периодическое изменение передвижений
- •Благодарности
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Свободнотекущие ритмы
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Захватывание искусственными принудителями
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Внутренняя десинхронизация и частичное захватывание
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Эксперименты со сдвигом фазы и перелеты в широтном направлении
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Приложения в области медицины
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Благодарность
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Рекомендуемая дополнительная литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Зависимость от времени дня
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Круглосуточные исследования
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Влияние сдвига фазы времязадателя
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Мотивация, обстановка и индивидуальные различия
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Заключительные замечания
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Литература
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Оглавление
- •Глава 1. Обзор биологических ритмов. Ю. Ашофф ... 12
- •Глава 2. Циркадианные системы: общая перспектива.
- •Глава 3. Свободнотекущие и захваченные циркадианные ритмы. Ю. Ашофф 54
- •Глава 4. Математические модели. Т. Павлидис .... 70
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Глава 5. Циркадианные системы: захватывание. К. Пит-
- •Глава 6. Ритмы поведения у беспозвоночных. Дж. Брэйди 12s
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
- •Глава 16. Ритмы работоспособности. П. Колькюхунь ... 389
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Уважаемый читатель!
- •Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с. Электронное оглавление
- •Глава 1. Обзор биологических ритмов 12
- •Глава 2. Циркадианные системы: общая перспектива 22
- •Глава 3. Свободнотекущие и захваченные циркадианные ритмы 54
- •Глава 4. Математические модели 70
- •Глава 5. Циркадианные системы: захватывание 87
- •Глава 6. Ритмы поведения у беспозвоночных 125
- •Глава 7. Нервный и эндокринный контроль циркадианной ритмичности у беспозвоночных 152
- •Глава 8. Генетика и развитие циркадианных ритмов беспозвоночных 188
- •Глава 9. Ритмы поведения позвоночных 200
- •Глава 10. Внутренняя временная упорядоченность 240
- •Глава 11. Нервный и эндокринный контроль циркадианных ритмов у позвоночных 275
- •Глава 12. Онтогенез циркадианных ритмов 292
- •Глава 13. Ежедневные приспособительные стратегии поведения 315
- •Глава 14. Ориентация в пространстве с помощью часов 348
- •Глава 15. Циркадианная система человека 362
- •Глава 16. Ритмы работоспособности 389
Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
Циркадданные системы: захватывание 115
|
|
|
Рис. 21. Оптимальные свойства колебателя ( и форма КСФ) для ночных и дневных видов (см. текст). В нижней части рисунка пунктиром указаны циркадианные фазы (У и B), на которые приходится соответственно утренний или вечерний световой импульс. Сплошные линии — фазы ('У и 'В) , в которые мгновенно переходит колебатель. Фаза 'B остается неизменной при разных фотопериодах, когда <T и Sy <SB. (По [21].) |
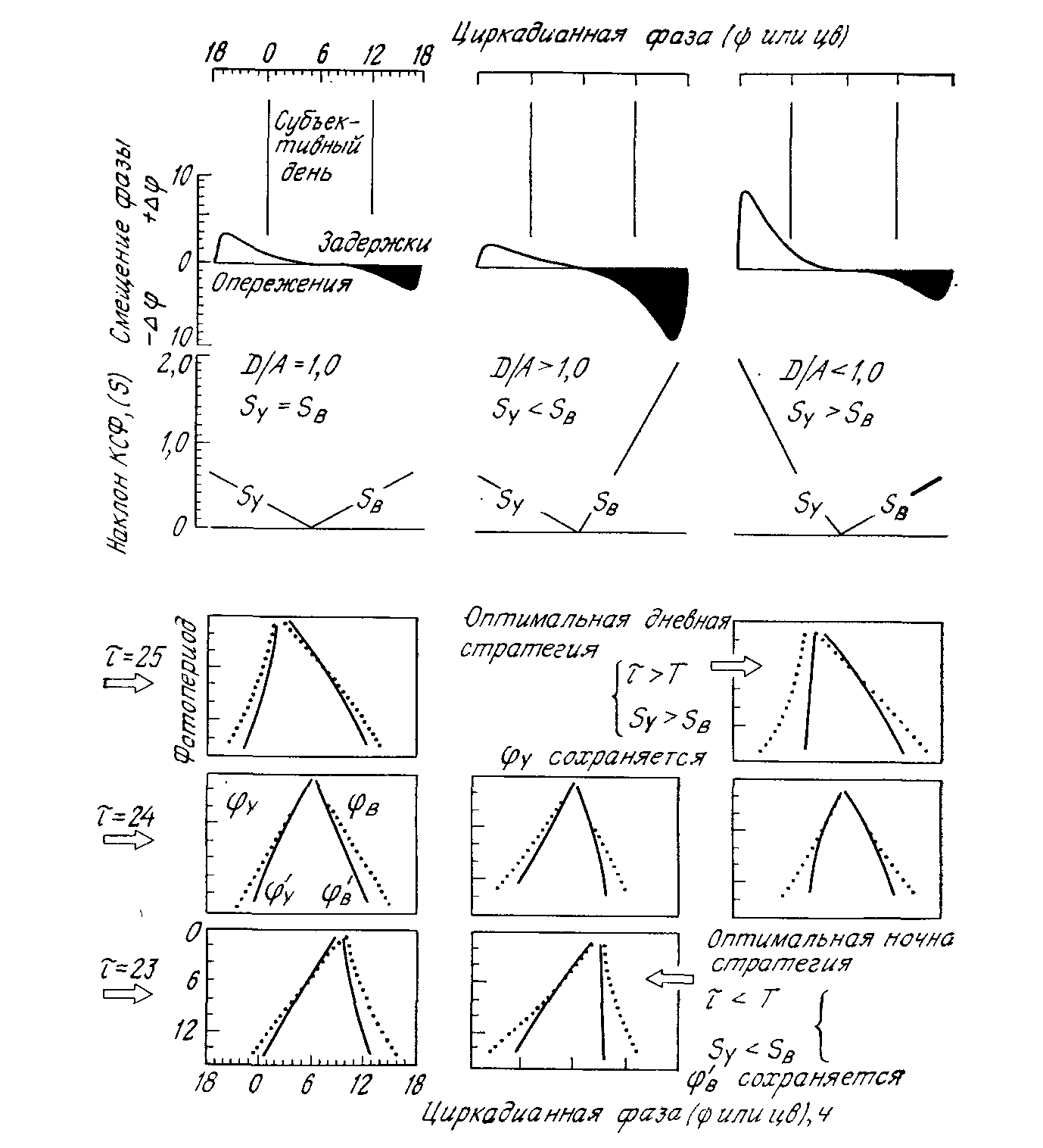
вания требует, чтобы суммарный суточный сдвиг фазы (У+ +B), вызываемый утренним и вечерним светом, оставался постоянным на протяжении всего года и был равен — 24 ч. Поэтому при увеличении продолжительности дня неизбежно некоторое сезонное смещение ночного колебателя относительно вечернего света, хотя оно может быть уменьшено путем увели-
Биологические ритмы. В 2-х т. Т. 1. Пер. С англ. — м.: Мир, 1984.— 414 с.
116 Глава 5
чения наклона SB вечернего участка КСФ в начале субъективной ночи (где происходит B) и уменьшения наклона Sy утреннего участка КСФ в конце субъективной ночи (где происходит У). Таким образом, всякое сезонное приращение ||, вызванное увеличением длины дня, ведет к уменьшению сезонного изменения B и (в том случае, если растущий наклон КСФ приближается к —1) практически возвращает колебатель к постоянному B. Тем самым сохраняется совпадение определенной фазы программы с закатом, но приносится в жертву совпадение другой фазы с рассветом. Из рис. 21 ясно, что с ростом —B в начале субъективной ночи одновременно растет наклон КСФ; значит, если уменьшается и —B, необходимое для захватывания, возрастает, то вечерний световой импульс приходится на ту часть КСФ, где наклон (SB) круче, и сезонные изменения B и 'B соответственно тоже уменьшаются.
Таким образом, при дискретном механизме захватывания ночные виды, для того чтобы сохранить постоянную фазу начала программы относительно вечернего импульса, должны иметь асимметричную КСФ с SB>SУ; кроме того, чем корочет, тем лучше будет сохраняться фаза . Для ночных грызунов и в самом деле характерны короткие и асимметричные КСФ с SB>SУ (и, следовательно, D>A) (см. рис. 8). Как отмечалось выше, существует эмпирическая корреляция между формой КСФ и величиной изменения периода под действием постоянного освещения (СС): одновременно с ростом D/A (а значит, и SB/SУ) возрастает , вызываемое режимом СС. Правило Ашоффа в отношении ночных видов гласит, что период y них обычно короче 24 ч и постоянный свет растягивает его больше, чем у дневных видов. Этих же особенностей (<24 ч; cc>>tt) можно ожидать и при использовании ночными видами оптимальной стратегии (<24 ч; SB>SУ, следовательно, D/A велико) в случае дискретного механизма захватывания. Очевидно, правило Ашоффа отражает широкую распространенность именно такой приспособительной стратегии у ночных животных [22, 24].
Если бы дневные виды поддавались захватыванию по тому же дискретному двухимпульсному механизму, правило Ашоффа и для них имело бы сходное функциональное значение. Оптимальная стратегия предполагала бы >24 ч и SУ>SB (следовательно, D/A мало); постоянный свет в этом случае не так сильно растягивал бы период колебателя, как у ночных видов (у которых D/A велико), и в крайних случаях (когда D>>A) вызывал бы даже уменьшение . Однако захватывание у дневных видов в природе осуществляется сложнее; наиболее очевидно то, что оно происходит не под действием дискретных утренних и вечерних импульсов. Полные фотопериоды, которые