

Метаболизм ксенобиотиков в организме
Большинство ксенобиотиков подвергается в организме биотрансформации. В неизменном виде выделяются высокогидрофильные ионизированные соединения. Из липофильных веществ исключение составляют средства для ингаляционного наркоза, основная часть которых не вступает в химические реакции в организме. Они выводятся легкими в том же виде, в каком были введены.
Метаболизм, или биологическая трансформация веществ, — понятие, включающее все химические изменения, происходящие с веществом в организме. В целом, все реакции биологической трансформации ксенобиотиков, в том числе, ЛС относят к одной из двух категорий, их обозначают как фазы метаболизма I и II. I фаза-модификация структуры ксенобиотика и II фаза - конъюгация ксенобиотика.
В результате метаболизма ксенобиотиков, с одной стороны, повышается их растворимость в воде, что способствует выведению из организма с мочой, а с другой — изменяется фармакологическая активность или токсичность данных веществ. В результате реакций метаболизма может происходить: а) полная потеря активности или токсичности вещества; б) активация или усиление токсичности в) изменение активности или появление нового токсического эффекта.
I фаза метаболизма ксенобиотиков
Метаболическая трансформация — I фаза метаболизма ксенобиотиков, включает несинтетические реакции, такие как окисление, восстановление, гидролиз. Метаболическая трансформация происходит за счёт присоединения или освобождения функциональных групп; гидроксильных (-ОН), сульфгидрильных (-SH) или аминогрупп (-NH2) в результате чего ксенобиотик становится более гидрофильным.
В метаболизме ксенобиотиков принимают участие ферменты почек, лёгких, кожи и ЖКТ, но наиболее активны ферменты печени.Основной вклад в обмен ксенобиотиков вносит эндоплазматическая сеть. Поскольку «микросомами» называютфракцию, полученную при дифференциальном центрифугировании клеточных гомогенатов и богатую эндоплазматической сетью, часто говорят о микросомальном и немикросомальных путях метаболизма. Вне микросом обмен может проходить в лизосомах, пероксисомах, митохондриях, цитозоле.
Микросомное окисление — совокупность реакций I фазы биотрансформации ксенобиотиков и эндогенных соединений, катализирующихся ферментными системами мембран эндоплазматического ретикулума гепатоцитов при участии цитохрома Р450 (рис. 29.1).
Микросомные ферменты катализируют реакции С-гидроксилирования, N-гидроксилирования, О-, N-, S-дезалкилирования, окислительного дезаминирования, сульфоокисления и эпоксидирования.
Цитохром Р450 (CYP) представляет группу ферментов (цитохром Р450-зависимые монооксигеназы), осуществляющих не только метаболизм ЛС и других ксенобиотиков, но и участвующих в синтезе глюкокортикостероидных гормонов, холестерола, желчных кислот, тромбоксанов и простациклинов. Субстратная специфичность этих ферментов очень низка, поэтому они окисляют различные вещества.
Цитохром Р450 — гемопротеин. Имеет множество изоферментов (более 1000 изоформ). Изоферменты цитохрома Р450 по идентичности аминокислотного состава разделяют на семейства (существует 17 семейств) и 39 подсемейств. Название цитохром Р450 указывает на то, что максимум поглощения комплекса цитохрома Р450 лежит в области 450 нм.
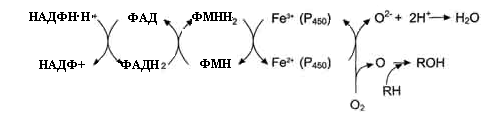
Рис. 30.1. Электронтранспортная цепь: НAДФH∙H+ ‒ P450 редуктаза ‒ цитохром Р450
В большинстве случаев донором электронов (е) для этой цепи служит НAДФH∙H+, окисляемый НAДФH.H+-P450 редуктазой. Фермент в качестве простетической группы содержит 2 кофермента ‒ флавинадениндинуклеотид (ФАД) и флавинмононуклеотид (ФМН). Протоны и электроны с НAДФH∙H+ переходят последовательно на коферменты. Восстановленный ФМН (ФМНH2) окисляется цитохромом Р450. Связывание в активном центре цитохрома Р450 вещества R-H активирует восстановление железа в геме ‒ присоединяется первый электрон. Изменение валентности железа увеличивает сродство комплекса P450-Fe2+·RH к молекуле кислорода. Появление в центре связывания цитохрома Р450 молекулы О2 ускоряет присоединение второго электрона и образование комплекса P450-Fe2+O2--RH. На следующем этапе Fe2+ окисляется, второй электрон присоединяется к молекуле кислорода P450-Fe3+O22-. Восстановленный атом кислорода О2- взаимодействует с протонами: О2- + 2Н+ → Н2О, и образуется вода. Второй атом молекулы кислорода включается в субстрат R-H, образуя гидроксильную группу вещества R-OH. Модифицированное вещество R-OH отделяется от фермента.
Таким образом, реакции с участием цитохромов Р450 заключаются в гидроксилировании веществ типа R-H с использованием одного атома молекулы кислорода О2, второй атом соединяется с протонами водорода H+ с образованием воды (поэтому ферменты называют также монооксигеназами или гидроксилазами).
Суммарное уравнение реакции гидроксилирования вещества R-H ферментами микросомального окисления:
R-H + О2 + НАДФН∙Н+ → R-OH + Н2О + НАДФ+
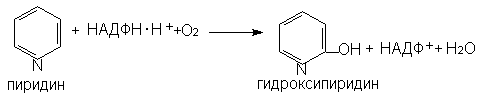
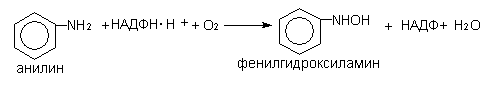
Гидроксилирование гетероциклических соединений
Гидроксилирование алифатических соединений
![]()
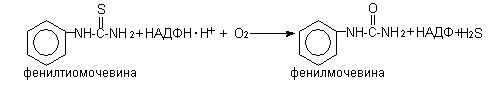
N-окисление с образованием N - оксидов и N – гидроксиламинов (аминазин, морфин, ацетиламинофлюорен).
S-окисление и десульфирование (аминазин, тиобарбитал).
Окислительное дезаминирование (амфетамин).
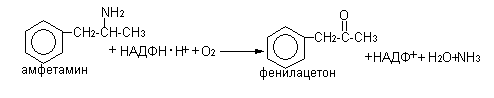
Удаление групп. Деалкилирование по N (морфин, лидокаин, атропин, диазепам).
RNHCH3 → RNH2 + H2C=O
Удаление групп. Деалкилирование по О (кодеин, фенацетин, кофеин, папаверин).
ROCH3 → ROH + H2CO
Дегалогенизация (хлороформ, метоксифлуран, галотан).
Эпоксидирование.
R-CH=CH-R'
→ R-CH=CH-R'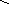

O
Метаболизм этанола.Цитохром Р450-зависимая микросомная этанолокисляющая система (МЭОС) локализована в мембране гладкого ЭПР гепатоцитов и играет незначительную роль в метаболизме небольших количеств алкоголя, но индуцируется этанолом, другими спиртами, барбитуратами и приобретает существенное значение при злоупотреблении этими веществами. При хроническом алкоголизме за счёт гипертрофии ЭПР и индукцииCYP2E1 (изофермент цитохрома Р450) окисление этанола ускоряется на 50‒70 %. Кроме того, этанол конкурирует с ксенобиотиками за связывание сCYP2E1, вызывая гиперчувствительность к некоторым принятым одновременно с ним лекарственным препаратам.
Кроме основной реакции, цитохром Р450катализирует образование активных форм кислорода (О2-, Н2О2), которые стимулируют перекисное окисление липидов.
Окисление немикросомными ферментами – дегидрогеназами и оксидазами.
Основную роль в метаболизме этанола и алифатических спиртов играет цинксодержащий НАД+- зависимый фермент — алкогольдегидрогеназа (АДГ), локализующаяся в основном в цитозоле и митохондриях печени (95%). АДГ катализирует обратимую реакцию, направление которой зависит от концентрации ацетальдегида и соотношения НАДФН∙Н+/НАД+в клетке.
С2Н5ОН + НАД+↔ СН3СНО + НАДФН∙Н+
Второстепенную роль в окислении этанола играет каталаза, находящаяся в пероксисомах цитоплазмы и митохондрий клеток печени. Этот фермент расщепляет примерно 2% этанола, но при этом утилизирует пероксид водорода.
СН3СН2ОН + Н2О2→ Н3СНО + 2 Н2О
Ацетальдегид, образовавшийся из этанола, окисляется до уксусной кислоты двумя ферментами: ФАД-зависимой альдегидоксидазой и НАД-зависимой ацетальдегиддегидрогеназой (АлДГ). Ацетальдегид - очень реакционно-способное соединение; он неферментативно может ацетилировать SH-, NH2- группа белков и других соединений в клетке и нарушать их функции.
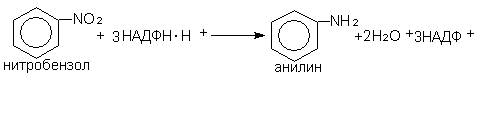
Восстановлению подвергаются отдельные ЛС (нитробензол, левомицетин, нитразепам, хлоралгидрат). Происходит это под действием нитроредуктаз и азоредуктаз.
восстановление нитросоединений
б) восстановление азосоединений
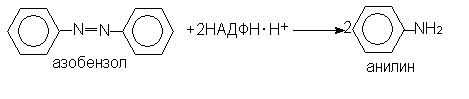
Гидролиз осуществляется в основном немикросомальными ферментами (эстеразами, амидазами, фосфатазами) в плазме крови и тканях. При этом вследствие присоединения воды происходит разрыв эфирных, амидных и фосфатных связей в молекулах. Гидролизу подвергаются сложные эфиры — ацетилхолин, новокаин, атропин, ацетилсалициловая кислота и амиды (новокаинамид).
Метаболиты, которые образуются в результате несинтетических реакций, могут в отдельных случаях обладать более высокой активностью, чем исходные соединения. Примером повышения активности в процессе метаболизма является использование предшественников лекарств — пролекарств, например салазопиридазин под действием фермента азоредуктазы кишечника превращается в сульфапиридазин и 5-аминосалициловую кислоту, обладающие антибактериальным и противовоспалительным действием (применяется для лечения язвенного колита). Химические превращения некоторых лекарств в организме приводят к изменению характера их активности. Например, ипразид — антидепрессант, в результате дезалкилирования превращается в изониазид, обладающий противотуберкулёзным действием.
Некоторые ксенобиотики в результате модификации структуры могут приобретать новые свойства и оказывать побочное действие на другие клетки (мутагенное, канцерогенное, иммунодепрессивное, аллергическое и т.д.).Так, эпоксиды, образовавшиеся при микросомальном окислении, являются канцерогенами. Они обладают высокой химической активностью и могут участвовать в реакциях неферментативного алкилирования ДНК, РНК, белков. Химические модификации этих молекул могут привести к перерождению нормальной клетки в опухолевую.
Однако, в большинстве случаев, образование в молекуле гидрофильных функциональных групп вызывает детоксификацию веществ и позволяет перейти процессу обезвреживания ко второй фазе — реакциям конъюгации.