

Фотосинтетические пигменты
У высших растений имеется три группы пигментов: хлорофиллы, каротиноиды и фикобилины.
Основными пигментами,
осуществляющими поглощение квантов
света в процессе фотосинтеза, являются
хлорофиллы, пигменты, содержащие
Mg-порфириновый комплекс. Обнаружено
несколько форм хлорофиллов, различающихся
по химическому строению. Спектр поглощения
различных форм хлорофиллов охватывает
видимую, ближнюю ультрафиолетовую и
ближнюю инфракрасную области спектра
(у высших растений от 350 до 700 нм, а у
бактерий — от 350 до 900 нм). Хлорофилл а
является основным пигментом и характерен
для всех организмов, осуществляющих
фотосинтез с выделением кислорода (рис
6.5). 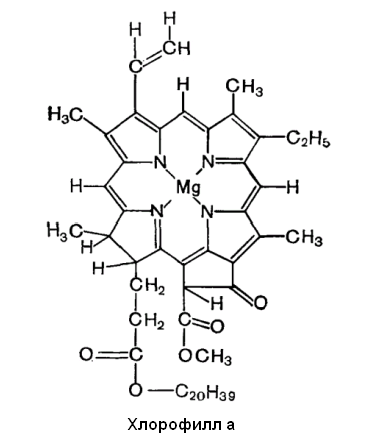
У фотосинтезирующих организмов кроме хлорофилла a имеются хлорофиллы b, с и d, которые расширяют спектр поглощения света.
Рис. 6.5. Строение хлорофилла а
В поглощении световой энергии участвуют каротиноиды (пигменты полиизопреноидной природы) — у фотосинтезирующих эукариот, и фикобилины (пигменты с открытой тетрапиррольной структурой) — у цианобактерий и красных водорослей.
В клетке молекулы хлорофилла вместе с другими пигментами, участвующими в процессах поглощения квантов света и передачи энергии, образуют светособирающие хлорофилл-белковые комплексы (ССК). Молекулы ССК имеют максимум поглощения при разной длине волны и расположены от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей.
Важнейшим структурно-функциональным звеном фотосинтетического аппарата является фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона.
В процессе фотосинтеза у растений принимают участие две фотосистемы.
Фотосистема I включает светособирающий комплекс и фотохимический реакционный центр I, в состав которого входит димер хлорофилла, поглощающий свет с длиной волны 700 нм (П700).
Фотосистема II включает светособирающий комплекс и фотохимический реакционный центр II, в состав которого входит димер хлорофилла, поглощающий свет с длиной волны 680 нм (П680).
Свет поглощается двумя фотосистемами раздельно, и нормальное осуществление фотосинтеза требует их одновременного участия.
Световая фаза фотосинтеза
Фотосинтез начинается с поглощения квантов света молекулами хлорофилла и другими связанными с ним пигментами. Энергия поглощенных квантов света стекается от сотен молекул пигментов ССК к молекуле пигмента П700 (Е0 = + 0,43В), которая переходит в возбужденное состояние (Е0 = ‒ 0,80 В) и легко отдает электрон первичному акцептору (фотохимическая реакция). Электрон с первичного акцептора, которым является мономерная форма хлорофилла а, передается на филлохинон (витамин К) – вторичный акцептор и затем на железосерные белки. Следующим переносчиком является железосодержащий белок ферредоксин (Е0 = ‒ 0,43В). Ферредоксин содержит два атома железа в негеминовой форме. От ферредоксина электрон переносится на НАДФ (Е0 = ‒ 0,32В). Этот перенос осуществляется с помощью специфического белка-фермента (ферредоксин-НАДФ-редуктазы), коферментом которого является ФАД.
Последовательность расположения переносчиков определяется величиной окислительно-восстановительного потенциала: электроны спонтанно текут в сторону менее отрицательного окислительно-восстановительного потенциала (рис. 6.6).

Рис 6.6. Электрон-транспортная цепь в мембране тилакоида
Отдав электрон, П700 остается в виде ионизированной молекулы. При этом потенциал П700становится снова + 0,43 В (основное состояние). Благодаря этому он является прекрасным акцептором электронов. Источником электрона, заполняющего эту «дырку», является фотосистема II. Она ответственна за реакции, связанные с разложением воды и выделением кислорода.
В состав реакционного центра фотосистемы II входит хлорофилл а, поглощающий свет с длиной волны 680 нм (П680). Под влиянием поглощенного кванта света возбужденный электрон от П680 (Е0 = ‒ 0,7 В) воспринимается первичным акцептором, которым является молекула феофитина. Затем электрон передается на пластохиноны, переносящие как электроны, так и протоны. От пластохинона электроны поступают на b/f-комплекс и передаются через железосерный белок на цитохром. Цитохром относится к группе цитохромов с (Е0 = +0 ,36 В). Воспринимая электрон, цитохром восстанавливается: Fe3+ + е- —> Fe2+. Следующий переносчик — пластоцианин — это медьсодержащий белок, в котором на каждую молекулу белка приходится два атома меди (Е0 = + 0,37 В), осуществляющих электронный транспорт: Cu2+ + е- —> Сu+. Пластоцианин выполняет роль связующего звена между b/f-комплексом и фотосистемой I. От пластоцианина электрон заполняет электронную «дырку» у П700.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав фотосистемы II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Водоокисляющий комплекс находится со стороны внутритилакоидного пространства, и полученные 4 протона выбрасываются внутрь тилакоида.
2Мn4+ + 2Н20 —> 2Мn2+ + 4Н+ + 4е- + 02
Таким образом, в результате работы фотосистемы II происходит окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве. Протоны внутрь тилакоида перекачиваются также через b/f-комплекс за счет энергии, которая выделяется в окислительно-восстановительной реакции при транспорте электронов. В результате на мембране тилакоида создается электрохимический потенциал, который является промежуточной формой запасания энергии и используется для синтеза АТФ протонной АТФ-синтазой.
Помимо полного нециклического пути переноса электрона, описанного выше, может протекать и циклический путь. В этом случае ферредоксин вместо НАДФ+ восстанавливает пластохинон, который переносит электрон назад на b/f-комплекс. В результате образуется бóльший протонный градиент и больше АТФ, но не восстанавливается НАДФ+.
Темновая фаза фотосинтеза протекает в строме и не является светозависимой.
С3-фотосинтез (цикл Кальвина, восстановительный пентозофосфатный цикл) состоит из трёх стадий (рис. 6.7):
карбоксилирование;
восстановление;
регенерация акцептора CO2.
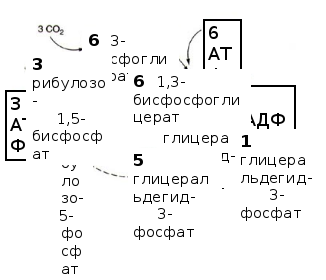
Рис. 6.7. Цикл Кальвина
На первой стадии к рибулозо-1,5-бисфосфату присоединяется CO2 под действием фермента рибулозобисфосфаткарбоксилазы.. Этот белок составляет основную фракцию белков хлоропласта и является наиболее распространённым ферментом в природе. В результате образуется промежуточное неустойчивое соединение (С6), распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК), которая является первичным продуктом фотосинтеза.
Во второй стадии ФГК фосфорилируется и восстанавливается с образованием глицеральдегид-3-фосфата (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 молекулы рибулозо-1,5-бисфосфата.
Две молекулы ФГА необходимы для синтеза глюкозы. Таким образом, для синтеза 1 молекулы глюкозы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН∙Н+ и 18 АТФ.
Интенсивность фотосинтеза зависит в первую очередь от интенсивности и спектрального состава света, концентрации СО2 и О2, температуры, водного режима растения, минерального питания и других факторов внешней среды.