

De novo синтез пиримидиновых нуклеотидов
В
отличие от пуринов, при биосинтезе
пиримидинов de novo(рис. 17.4) вначале образуется пиримидиновое
кольцо, а лишь затем к нему присоединяется
рибозо-5-фосфат.
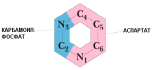
Источниками атомов для пиримидинового
кольца являются глутамин,
аспартат и СО2.
Синтез начинается с образования
карбамоилфосфатапод действием
карбамоилфосфатсинтетазыII(КФС II). Отличия
отКФС I:

|
КФС I |
КФС II |
|
|

Рис. 17.4. De novo синтез пиримидиновых нуклеотидов
У млекопитающих первые три реакции процесса катализирует полифункциональный фермент, который, помимо карбамоилфосфатсинтетазного, содержит аспартаткарбамоилтрансферазный и дигидрооротазный каталитические центры; оротатфосфорибозилтрансфераза и ОМФ-декарбоксилаза также являются составляющими одного белка – УМФ-синтазы. Ключевым этапом в синтезе пиримидиновых нуклеотидов у млекопитающих считается образование карбамоилфосфата.
Регуляция: процесс ингибируется избытком пиримидиновых нуклеотидов, а избытком пуриновых и ФРПФ — активируется.
Оротовая ацидурия— причиной этого заболевания является дефект ферментов, превращающих оротовую кислоту в УМФ. Характерно отставание в физическом и умственном развитии, мегалобластическая анемия. В лечении используют уридин.
Образование дезоксирибонуклеотидов
Образование дезоксирибонуклеотидов, необходимых для биосинтеза ДНК, происходит на уровне нуклеозиддифосфатов (АДФ, ГДФ, ЦДФ, УДФ) с участием специального фермента (редуктаза) и белка тиоредоксина (рис. 17.5). Тиоредоксин содержит свободные SH-группы, которые являются донорами атомов водорода для реакции восстановления 2/-ОН группы в рибозе. В результате реакции рибоза в составе НДФ превращается в дезоксирибозу. Затем дНДФ превращаются в дНТФ и используются для синтеза ДНК.
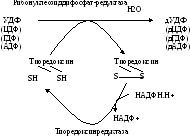
Рис. 17.5. Образование дезоксирибонуклеотидов
Тема 18. Биосинтез днк, рнк и белка
Центральная догма молекулярной биологии отражает поток информации в клетке:
ДНК
РНК БЕЛОК


Биосинтез днк
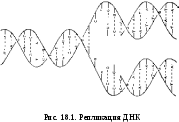
Репликация — процесс удвоения ДНК (синтез ДНК на матрице ДНК) (рис. 18.1).
Принципы репликации:
1) комплементарность;
2) антипараллельность;
3) однонаправленность;
4) потребность в праймере (затравке);
5) прерывистость;
6) полуконсервативность.
Первые 3 принципа можно сформулировать одной фразой: синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи и всегда в направлении 5/ 3/.
Ферменты и белки, участвующие в репликации (их > 40), объединены в единый комплекс — реплисому.
Хеликаза — раскручивает двойную спираль ДНК в репликационной вилке.
Топоизомераза — снимает напряжение, возникающее в репликационной вилке, и предотвращает обратное скручивание цепей.
Праймаза — синтезирует праймеры. Праймаза является РНК-полимеразой, поэтому образующиеся праймеры представляют собой олигорибонуклеотиды.
ДНК-полимераза — главный фермент процесса. Компоненты, необходимые для её работы: матрица, затравочный олигонуклеотид (праймер), субстраты (активированные нуклеотиды — дАТФ, дГТФ, дЦТФ, дТТФ), ионы магния. ДНК-полимераза катализирует реакцию образования фосфодиэфирной связи с использованием энергии макроэргического субстрата:
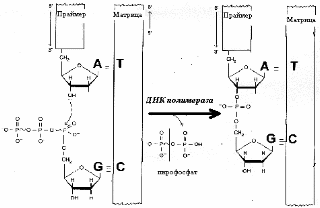
Большинство ДНК-полимераз обладают способностью исправлять ошибки, допущенные при синтезе, путем отщепления неправильно присоединенного нуклеотида и замены его на нужный.
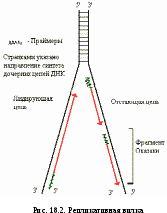
Поскольку цепи ДНК антипараллельны, а синтез идет только от 5/-конца к 3/-концу, одна из дочерних цепей синтезируется прерывисто, образуются фрагменты Оказаки (рис. 18.2). Впоследствии праймеры (участки РНК) из дочерней цепи удаляются, на их месте достраивается ДНК.
ДНК-лигаза — сшивает фрагменты, образующиеся после удаления праймеров и достройки ДНК.
После окончания репликации ДНК подвергается метилированию (защита от нуклеаз).
У прокариот есть
три ДНК-полимеразы – ДНК-полимераза
III
(непосредственно ведет репликацию),
ДНК-полимераза II
(участвует в репарации), ДНК-полимераза
I
(отвечает за удаление праймеров и
достройку на их месте ДНК).У
эукариот одновременно с репликацией
идет синтез гистонов. Ферменты:
ДНК-полимераза α (отвечает за инициацию
процесса и синтез праймеров), β
(репаративная), γ (митохондриальная), δ
(синтезирует лидирующую цепь), ε
(наращивает отстающую цепь). На концах
линейных хромосом эукариот имеются
теломеры (неинформативные повторяющиеся
последовательности нуклеотидов). В
соматических клетках с каждым актом
репликации теломеры укорачиваются
из-за невозможности достроить ДНК на
месте 5/-праймера.
Это своеобразные «молекулярные часы»
клетки.
Стабильность и неизменность структуры ДНК обеспечивается: 1) высокой точностью репликации; 2) специальными механизмами исправления повреждений, возникших в ДНК, - это пострепликационная репарация. Возможна прямая репарация (химические реакции, восстанавливающие исходную структуру поврежденного нуклеотида) и эксцизионная репарация (вырезание поврежденного нуклеотида или азотистого основания с последующим заполнением дефекта) (рис. 18.3).
В природе существует путь синтеза ДНК на матрице РНК с участием обратной транскриптазы (или РНК-зависимой ДНК-полимеразы). Благодаря наличию этого фермента некоторые РНК–содержащие вирусы имеют возможность переписывать свою генетическую информацию на ДНК клеток хозяина.
Рис. 18.3. Эксцизионная
репарация