

Синтез жирных кислот
Субстраты синтеза жирных кислот. Предшественником является ацетил-КоА, процесс протекает в цитозоле (рис. 12.3). Ацетил-КоА образуется из пирувата под действием митохондриального пируватдегидрогеназного комплекса. Внутренняя митохондриальная мембрана непроницаема для ацетил-КоА. В митохондриях фермент цитратсинтаза катализирует реакцию образования цитрата из ацетил-КоА и ЩУК. Цитрат выходит из митохондрий в цитоплазму. В цитозоле фермент АТФ-цитратлиаза расщепляет цитрат до ацетил-КоА и ЩУК.

Рис. 12.3. Субстраты для синтеза жирных кислот. Переход ацетил-КоА из митохондрий в цитозоль
Ферменты синтеза жирных кислот. В биосинтезе насыщенных жирных кислот участвуют два ферментных комплекса: ацетил-КоА карбоксилаза и ацилсинтетаза (рис. 12.4).
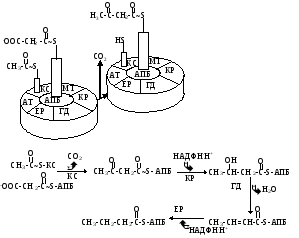
Рис. 12.4. Схема реакций синтеза жирных кислот
Ацилсинтетазный мультиферментный комплекс. Комплекс содержит ацилпереносящий белок (АПБ) в качестве своеобразного ядра. Другими ферментами комплекса являются -кетоацилсинтетаза (КС), малонилтрансфераза (МТ), -кетоацилредуктаза (КР), -гидро-ксиацилдегидратаза (ГД), еноилредуктаза (ЕР) и ацилтрансацетилаза (АТ).
Ацетил-КоА карбоксилаза катализирует ключевую реакцию в синтезе жирных кислот:

Цитрат активирует фермент. Ацетил-КоА карбоксилаза подвергается обратимому фосфорилированию/дефосфорилированию; цАМФ-зависимая протеинкиназа ингибирует ферментативную активность, а фосфатаза — активирует. На этом основана гормональная регуляция активности ацетил-КоА карбоксилазы. Повышенные концентрации малонил-КоА и ацил-КоА аллостерически ингибируют активность этого фермента.
За 7 таких циклов образуется насыщенная жирная кислота с 16 углеродными атомами. В общем виде это выглядит следующим образом:
ацетил-КоА + 7 малонил-КоА + 14 НАДФН.Н+ + 14 Н+ пальмитат + 7 СО2 + 14 НАДФ + 8 КоА-SH + 6 Н2О
Потребность в НАДФН.Н+ для реакций восстановления в процессе синтеза жирных кислот. Источником восстановленного НАДФ+ является окислительный этап пентозофосфатного пути обмена глюкозы, а также реакции переноса водорода в пути расщепления цитрата. Последние протекают с участием фермента малатдегидрогеназы (кофермент НАДН.Н+) и малик-фермента (в реакции восстановления НАДФ+).
ПРОИСХОЖДЕНИЕ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ В КЛЕТКАХ
Незаменимые и заменимые. В организме человека не могут синтезироваться -3 и -6 жирные кислоты в связи с отсутствием ферментной системы, которая могла бы катализировать образование двойной связи в положении -6 или любом другом положении, близко расположенном к -концу. К таким жирным кислотам относятся: линолевая кислота (18:2, 9, 12), линоленовая кислота (18:3, 9, 12, 15), арахидоновая кислота (20:4, 5, 8, 11, 14). Арахидоновая кислота является незаменимой только при недостатке линолевой кислоты.
Функции полиненасыщенных жирных кислот: придают жидкостность мембранам, являются предшественниками эйкозаноидов
Эйкозаноиды
(липидные гормоны). Простагландины,
тромбоксаны и лейкотриены. Самым главным
предшественником является арахидоновая
кислота.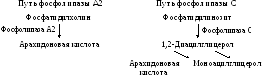
Схема высвобождения арахидоновой кислоты из мембранных фосфолипидов:
Судьба арахидоновой кислоты, которая высвободилась из состава мембранных фосфолипидов:
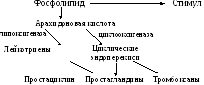
Ингибиторы синтеза эйкозаноидов. Кортикостероиды ингибируют фосфолипазу А2, тем самым они ингибируют высвобождение арахидоновой кислоты из состава мембранных фосфолипидов. Ингибиторы из числа противовоспалительных препаратов нестероидной природы (аспирин, индометацин, фенилбутазон) ингибируют циклоксигеназу.
Простагландины стимулируют сокращение гладких мышц, липолиз, снижают секрецию желудочного сока, свертывание крови, вызывают расширение кровеносных сосудов.
Тромбоксаны стимулируют агрегацию тромбоцитов, сокращение стенки артерий, образование цАМФ в тромбоцитах, повышают кровяное давление.
Лейкотриены участвуют в формировании воспаления, аллергических реакций, в хемотаксисе.