

Процессинг препроколлагена
Полипептидные цепи синтезируются на полисомах в виде препроколлагена («пре» указывает на наличие сигнального, лидирующего пептида; «про» — на наличие дополнительных пептидов на N- и С-концах, которые необходимы для формирования правильной пространственной структуры). Затем начинается процессинг препроколлагена (рис. 27.2), в котором выделяют внутриклеточный и внеклеточный этапы.
Внутриклеточнопроисходит удаление сигнального пептида и проколлаген поступает в ЭПР.
Полипептидные цепи гидроксилируются пролилгидроксилазой и лизингидроксилазой. Для работы этих ферментов в качестве кофакторов необходимы О2, Fe2+, α-кетоглутарат и аскорбиновая кислота. Снижение содержания гидроксипролина клинически проявляется цингой (при дефиците аскорбиновой кислоты), которая является следствием нестабильности молекул из-за изменения их структуры и разрушения протеазами.
Происходит гликозилирование остатков гидроксилизина, что необходимо для секреции молекул проколлагена в межклеточное пространство.
В дополнительных пептидах на N- и С-концах формируются межцепочечные дисульфидные связи между радикалами цистеина, что сопровождается образованием тройной спирали коллагена.
Секреция
проколлагена во внеклеточное пространство.
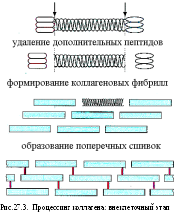
Ферменты амино- и карбоксипептидазы удаляют дополнительные пептиды. После чего становится возможной организация молекул коллагена в фибриллы (микрофибриллы) и переплетение фибрилл с образованием коллагеновых волокон (рис.27.3).
Оксидазы (для них необходимы Cu2+ и витамин.B6) катализируют окислительное дезаминирование ε-аминогрупп лизина и гидроксилизина в молекулах коллагена с образованием альдегидных групп. Конденсация альдегидных групп сопровождается образованием поперечных связей в коллагеновых волокнах.
Эластин
Эластин — самый прочный из белков, известных в организме человека.
В отличие от коллагена у эластина один генетический тип, мало ОН-ПРО, нет ОН-ЛИЗ, дополнительных пептидов, углеводов, не образуется тройная спираль.
Молекула эластина
состоит из двух типов фрагментов,
чередующихся вдоль цепи: гидрофобные
(фибриллярные) сегменты, которые
ответственны за эластические свойства
молекулы и глобулярные сегменты, богатые
АЛА и ЛИЗ, имеющие форму α-спирали и
участвующие в формировании поперечных
связей между молекулами эластина.
Синтезируется эластин в виде мономера,
а внеклеточно происходит фибриллогенез
с образованием поперечных связей. В
образовании поперечных связей принимаю
участие 4 остатка ЛИЗ. С участием оксидаз
происходит окислительное дезаминирование
лизина и последующая межмолекулярная
конденсация 4-х альдегидных групп. В
результате чего образуется особый вид
межмолекулярных сшивок – десмозин
(рис. 27.4) или лизиннорлейцин,
которые присущи только эластину.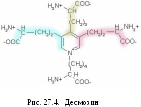
Фибриллярные адгезивные белки
Внеклеточный матрикс содержит большое число адгезивных неколлагеновых белков, структурной особенностью которых является наличие доменов, способных специфически связываться с другими макромолекулами и рецепторами на поверхности клетки. Непременным компонентом доменов, обеспечивающих взаимодействие с клетками, является последовательность аминокислот АРГ-ГЛИ-АСП (R-G-D).
Фибронектин — высокомолекулярный гликопротеин. Существуют множественные формы фибронектина. Одна из них — фибронектин плазмы и других биологических жидкостей. Он принимает участие в механизмах свертывания крови и заживления ран. Фибронектины тканей располагаются на поверхности клеток, образуя фибронектиновые филаменты.
Фибронектин ускоряет клеточную миграцию, обеспечивая взаимодействие клеток с матриксом.
Фибриллин — структурный компонент микрофибрилл, обеспечивающих образование эластиновых волокон. Он найден в хрусталике, периосте, аорте. При мутации гена, кодирующего синтез фибриллина, развивается синдром Марфана: эктопия хрусталика, арахнодактилия («паучьи» пальцы), поражение суставов.
Ламинин и энтактин — гликопротеины базальной мембраны. Они связываются не только между собой, но и с изоколлагеном IV , гепарансульфатом, поверхностью эпителиальных клеток, причем для связывания с различными веществами имеются свои домены.
Каждый тип соединительной ткани имеет свои специфические наборы молекул: кроме соответствующих изоколлагенов, имеются и специфические неколлагеновые белки.
В хрящевой: главный ПГ и минорные ПГ (фибромодулин — регулятор фибриллогенеза; бигликан — значение его пока неизвестно; декорин — способен связываться с изоколлагеном II и играет роль ингибитора фибринолиза; белки с разной молекулярной массой и не очень изученными функциями, из известных функций — связывание с хондроцитами, кристаллами гидроксиапатита, изоколлагеном II для его фиксации к хондроцитам).
В костной: индукторы и ингибиторы остеогенеза, инициаторы минерализации. Это неколлагеновые белки.
Остеокальцин: 1) содержит 3 остатка γ-карбокисиглутаминовой кислоты → связывает Са2+ ; 2) прочно связан с апатитом; 3) участвует в росте кристаллов.
Костный сиалопротеин: 1) содержит трипептид АРГ-ГЛИ-АСП (R-G-D) → способен связываться с другими клетками, макромолекулами и рецепторами клеточных мембран; 2) через специальный рецептор (10 остатков ГЛУ) связывается с Са2+; 3) относится к фосфопротеинам, тесно связан с клетками и апатитом; 4) присоединяет остеобласты к кости в период ее синтеза.
Остеопонтин: 1) содержит трипептид R-G-D; 2) связывается с Са2+; 3) прочно связан с апатитом.
Остеонектин: 1) имеет Са-связывающий домен, хотя в нем и нет γ-карбокисиглутаминовой кислоты; 2) связывается с коллагеном и апатитом.
Тромбоспондин: 1) содержит трипептид R-G-D; 2) связывается с поверхностями клеток и другими белками костной ткани.
Костный кислый гликопротеин: участвует в минерализации костной ткани.