

Патогенетическая взаимосвязь углеводов пищи и кариеса
Потребление легкоферментируемых углеводов, в частности сахарозы, инициирует кислотную деминерализацию эмали зубов. У бактерий имеются два альтернативных пути использования пирувата: первый — путь восстановления ПВК в молочную кислоту с участием лактатдегидрогеназы (ЛДГ), второй — расщепление ПВК на уксусную и муравьиную кислоту с участием пируватформиатлиазы (ПФЛ) (рис. 8.3).

Уксусная кислота + Муравьиная кислота
Рис. 8.3 Схема расщепления сахарозы под влиянием ферментов бактерий полости рта
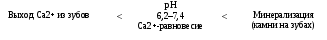
Существует определенная зависимость между рН ротовой жидкости и выходом кальция из зубов:
Аэробное окисление глюкозы
Это основной путь катаболизма глюкозы у аэробных организмов. Процесс осуществляется в три этапа (рис. 8.4). В аэробных условиях глюкоза окисляется до СО2 и Н2О.
Рис. 8.4.
Этапы аэробного окисления глюкозы
Энергетический баланс. Энергетический баланс аэробного окисления глюкозы — 30 – 32 моля АТФ на молекулу глюкозы
Челночные механизмы переноса восстановительных эквивалентов через митохондриальные мембраны. В аэробных условиях окисление гликолитического восстановленного никотинамидадениндинуклеотида (НАДН.Н+) осуществляется в митохондриях в ходе тканевого дыхания. Цитозольный НАДН.Н+ не может передавать водород непосредственно на дыхательную цепь, поскольку внутренняя мембрана митохондрий непроницаема для НАДН.Н+ . Перенос водорода с цитозольного НАДН.Н+ в митохондрии происходит при участии специальных механизмов, называемых челночными (рис. 8.5). Суть этих механизмов состоит в том, что НАДН.Н+ в цитозоле восстанавливает некоторое соединение, способное проникать в митохондрию; в митохондрии это соединение окисляется, восстанавливая при этом внутримитохондриальный НАД (малатный челночный механизм) или ФАД (глицеролфосфатный челночный механизм), и вновь переходит в цитозоль.
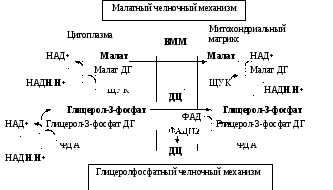
Рис. 8.5. Челночные механизмы транспорта цитоплазматического НАДН.Н+
Глюконеогенез
Глюконеогенез — синтез глюкозы из соединений неуглеводной природы.
В организме взрослого человека за сутки может синтезироваться до 250 г глюкозы.
Глюконеогенез осуществляется главным образом в печени (синтезируетя до 90 % всей глюкозы), корковом веществе почек, энтероцитах (совсем незначительно).
Глюконеогенез стимулируется при длительном голодании, при ограничении поступления углеводов с пищей, в период восстановления после мышечной нагрузки, у новорождённых в первые часы после рождения.
Субстраты глюконеогенеза. Истинными субстратами глюконеогенеза являютя пируват, оксалоацетат, фосфодиоксиацетон, которые непосредственно включаются в этот процесс. Все вещества неуглеводной природы, дающие эти метаболиты, являются субстратами глюконеогенеза: лактат→ПВК, метаболиты цикла Кребса→ЩУК, глицерол→фосфодиоксиацетон, пропионил-КоА→метаболиты цикла Кребса→ЩУК, глюкогенные аминокислоты→ПВК или ЩУК. Главный источник субстратов глюконеогенеза — глюкогенные аминокислоты. К глюкогенным аминокислотам относятся все протеиногенные аминокислоты, кроме лейцина и лизина.
Стехиометрия:
2ПВК + 4АТФ + 2ГТФ + 2НАДН.Н+ + 6Н2О ® Глюкоза + 4АДФ + 2ГДФ + 6Фн + 2НАД +
Глюконеогенез протекает, в основном, по тому же пути, что и гликолиз, но в обратном направлении. Для обхода трех ключевых реакций гликолиза используются четыре специфических фермента глюконеогенеза.
1. Пируваткарбоксилаза
СН3 СООН
½ Биотин ½
С=О + СО2
+ АТФ СН2
+ АДФ + ФН
½ Å ½
С ООН Ацетил-КоА С=О
Пируват ½ Оксалоацетат
СООН
2. Фосфоенолпируваткарбоксикиназа
СООН
½
СН2 + ГТФ
½
С=О
СО2
½
Фосфоенолпируват
СООН
Оксалоацетат
3. Фруктозо-1,6-дифосфатаза
Фруктозо-1,6-дифосфат + Н2О Фруктозо-6-фосфат + ФН
4. Глюкозо-6-фосфатаза
Глюкозо-6-фосфат + Н2О Глюкоза + ФН
Рис. 8.6. Ключевые ферменты и ключевые реакции глюконеогенеза:
Энергетический баланс. На синтез молекулы глюкозы из двух молекул пирувата расходуется 4АТФ и 2ГТФ (6АТФ). Энергию для глюконеогенеза поставляет процесс β-окисления жирных кислот.
Регуляция глюконеогенеза. Глюконеогенез стимулируется в условиях гипогликемии при низком уровне инсулина и преобладании его антагонистов (глюкагона, катехоламинов, глюкокортикоидов).
1. Регуляция активности ключевых ферментов:
- фруктозо-1,6-дифосфатаза по аллостерическому механизму активируется АТФ, ингибируется фруктозо-2,6-дифосфатом и АМФ;
- пируваткарбоксилаза активируется СН3СО~SКоА (аллостерический активатор).
2. Регуляция количества ключевых ферментов: глюкокортикоиды и глюкагон индуцируют синтез ключевых ферментов, а инсулин — репрессирует.
3. Регуляция количества субстрата: количество субстратов глюконеогенеза увеличивается под действием глюкокортикоидов (катаболическое действие на белки мышечной и лимфоидной ткани, на жировую ткань), а также глюкагона (катаболическое действие на жировую ткань).
Биологическая роль глюконеогенеза:
1. Поддержание уровня глюкозы в крови. При длительном голодании (голодание более суток) глюконеогенез является единственным процессом, поставляющим глюкозу в кровь.
2. Возвращение лактата в метаболический фонд углеводов. Лактат, образующийся в процессе анаэробного окисления глюкозы в эритроцитах и скелетных мышцах, транспортируется кровью в печень и превращается в гепатоцитах в глюкозу. Это так называемый межорганный цикл Кори (рис. 8.7).
3. Предотвращение лактатного ацидоза, то есть в ходе глюконеогенеза лактат крови превращается в глюкозу.

Рис. 8.7. Цикл Кори