

Тема 21. Биохимия печени
Печень играет центральную роль в обмене веществ. У взрослого человека масса печени составляет ≈ 1,5 кг, при этом более 70 % приходится на долю воды. Из сухого остатка свыше 50 % составляют белки, 90 % из них — глобулины. В печени содержатся 100-150 г гликогена и многие ферменты. Около 5 % от массы печени составляют липиды (триацилглицеролы, фосфолипиды, холестерол),
Функции печени
1. Гомеостатическая. Печень поддерживает постоянный состав крови (гомеостаз). Всасывание питательных веществ из желудочно-кишечном тракте в кровь происходит непостоянно, в связи с чем в портальном круге кровообращения могут наблюдаться изменения концентраций веществ в разные периоды времени, однако благодаря гомеостатической функции печени концентрации важнейших метаболитов (глюкозы, аминокислот и др.) в большом круге кровообращения практически постоянны.
2. Роль печени в обмене углеводов. Основная роль печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это достигается благодаря регуляции процессов утилизации и образования глюкозы в печени, таких как:
синтез и распад гликогена;
глюконеогенез (синтез глюкозы из лактата, глицерола, аминокислот и других неуглеводных источников);
превращение гексоз (галактозы и фруктозы) в глюкозу;
пентозофосфатный путь;
образование и использование глюкуроновой кислоты.
3. Роль печени в обмене белков. Освобождающиеся в процессе пищеварения аминокислоты попадают с током крови в печень и используются для:
синтеза большинства белков плазмы: альбуминов, глобулинов, транспортных, свертывающей системы и фибринолиза;
образования α-кетокислот путём трансаминирования и окислительного дезаминирования аминокислот;
глюконеогенеза (гликогенные аминокислоты);
синтеза кетоновых тел (кетогенные и смешанные аминокислоты);
получения энергии, подвергаясь превращениям в цикле трикарбоновых кислот;
синтеза мочевины;
синтеза мочевой кислоты;
синтеза креатина, холина.
4. Роль печени в обмене липидов. Печень играет ключевую роль в метаболизме липидов, так как в печени:
синтезируются, окисляются, удлиняются и укорачиваются жирные кислоты, поступающие с пищей или образующиеся при распаде простых и сложных липидов;
синтезируются и распадаются триацилглицеролы и фосфолипиды;
синтезируются ЛПОНП и ЛПВП;
метаболизм остатков хиломикронов, ЛПОНП, ЛПНП и ЛПВП;
синтезируется холестерол (90 % из общего количества в организме);
синтезируются желчные кислоты, которые входят в состав желчи, необходимой для переваривания липидов в кишечнике;
синтезируются кетоновые тела (только в печени!).
5. Роль печени в обмене витаминов.
всасывание жирорастворимых витаминов (желчные кислоты), их депонирование и транспорт (липопротеины);
активация витаминов: каротин → витамин А, холекальциферол → 25-гидроксихолекальциферол (предшественник витамина D3);
депонирование витамина В12.
5. Антитоксическая функция. Поступающие в организм ксенобиотики (чужеродные вещества, в том числе лекарственные средства) и образующиеся в самом организме токсичные или непригодные для дальнейших превращений продукты метаболизма обезвреживаются в печени. 1
6. Роль печени в обмене желчных пигментов. Продолжительность жизни эритроцитов составляет приблизительно 120 дней, после чего происходит их разрушение и освобождение гемоглобина. При разрушении гемоглобина его белковая часть (глобин) гидролизуется до аминокислот. Порфириновая часть гема (без железа) деградирует до образования желчных пигментов, которые выводятся из организма. Главным органами, в которых осуществляется разрушение эритроцитов, является печень, селезёнка и костный мозг. Распад гемоглобина протекает в макрофагах, в звёздчатых ретикулоэндотелиоцитах, а также в гистиоцитах соединительной ткани по следующей схеме (рис. 21.1):
НАДФН∙Н+;
О2 глобин
НАДФН∙Н+
гемоглобинвердоглобинбиливердинбилирубин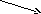
вит. С; цит. Р450 Fe биливердинредуктаза
гемоксигеназа
Рис. 21.1. Распад гемоглобина
Распад гемоглобина протекает с образованием гемоглобино-гаптоглобинового комплекса, который подвергается ферментативному окислению. Первая реакция катаболизма гема происходит при участии НАДФH-зависимого ферментативного комплекса гемоксигеназы (гем индуцирует транскрипцию гена гемоксигеназы). Ферментная сисгема локализована в мембране ЭПР. В результате образуется вердоглобин (пигмент зеленого цвета). От вердоглобина освобождается атом железа и белок и образуется линейный тетрапиррол — биливердин (пигмент зеленого цвета) и монооксид углерода. Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина НАДФН-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, но также при катаболизме других гемсодержащих белков, таких как цитохромы и миоглобин.Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), не растворяется в воде, токсичен, не проходит через почечные мембраны. Он составляет около 75% общего билирубина крови, по крови транспортируется в комплексе с альбумином. Этот билирубин называют непрямым (он даёт непрямую реакцию с диазореактивом Эрлиха), а также свободным, или неконьюгированным. Каждая молекула альбумина связывает 3 молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге рН крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за высокоаффинный, имеющий высокое сродство центр альбумина. Дальнейший метаболизм билирубина осуществляется в печени, где происходит процесс соединения (коньюгация) непрямого билирубина с 2 молекулами глюкуроновой кислотой при участии фермента УДФ-глюкуронилтрансферазы. Донором глюкуроновой кислоты является УДФ-глюкуронат. Индуктором синтеза УДФ-глюкуронилтрансферазы служит фенобарбитал. Образовавшийся билирубиндиглюкуронид называется прямой, связанный, или конъюгированный билирубин. Он не токсичен, растворяется в воде, проходит через почечный барьер, дает прямую реакцию с диазореактивом. В норме в крови его содержание не более 25% от общего билирубина. Секреция конъюгированного билирубина в жёлчь идёт по механизму активного транспорта, т.е. против градиента концентрации. Поступившие с желчью билирубинглюкурониды в кишечнике гидролизуются специфическими бактериальными ферментами β-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты. Освободившийся в ходе этой реакции билирубин восстанавливается с образованием мезобилирубина. Последний в дистальных отделах тонкого кишечника под действием кишечной микрофлоры восстанавливается с образованием уробилиногена (бесцветное тетрапиррольное соединение). Часть уробилиногена в тонкой кишке всасывается и через систему воротной вены переносится в печень, расщепляясь там до дипирролов. Основное количество уробилиногена в толстом кишечнике при участии анаэробной микрофлоры восстанавливается до стеркобилиногена. 80% стеркобилиногена выделяется с калом, окисляясь кислородом воздуха с образованием стеркобилина, придающего характерную окраску стулу. Часть стеркобилиногена адсорбируется в средней и нижней геморроидальных венах, поступает в систему нижней полой вены, а затем выводится с мочой. В норме уробилиноген в общий кровоток не поступает и с мочой не выделяется.
Норма общего билирубина в крови — 8,55–20,52 мкмоль/л, из них более 75 % приходится на долю свободного билирубина. При повышении концентрации общего билирубина в крови более 25 мкмоль/л у человека желтеют кожные покровы, слизистые оболочки и склеры. Такое состояние называется «желтуха» (icterus). Сведения о видах желтух и их дифференциальной диагностике представлены в таблице 21.1.
Таблица 21.1