

Тема 20. Гормоны. Общий механизм действия гормонов
Гормоны — это класс регуляторных молекул, синтезируемых специальными клетками.
Классификация гормонов по химической структуре:
Производные аминокислот: производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин; производные триптофана: мелатонин, серотонин; производные гистидина: гистамин.
Белково-пептидные гормоны: полипептиды: глюкагон, кортикотропин, меланотропин, вазопрессин, окситоцин, пептидные гормоны желудка и кишечника; простые белки: инсулин, соматотропин, пролактин, паратгормон, кальцитонин; сложные белки (гликопротеины): тиреотропин, фоллитропин, лютропин.
Стероидные гормоны: кортикостероиды (альдостерон, кортизол, кортикостерон); половые гормоны: андрогены (тестостерон), эстрогены и прогестерон.
Производные жирных кислот: арахидоновая кислота и ее производные: простагландины, простациклины, тромбоксаны, лейкотриены.
Классификация гормонов по механизму действия:
Взаимодействующие с рецепторами на поверхности мембраны (пептидные гормоны, адреналин, эйкозаноиды);
Гормоны, взаимодействующие с внутриклеточными рецепторами (стероидные и тиреоидные гормоны).
Для гормонов характерны следующие особенности биологического действия:
низкая концентрация в крови (10–6–10–12 М);
регуляция секреции по принципу прямой и (или) обратной связи;
обязательная, высокоспецифичная связь с рецептором, включающим каскадный механизм усиления гормонального сигнала;
реализация клеточного ответа путем изменения количества или активности ферментов.
Гормональный сигнал способен «выключаться» в результате инактивирования рецептора путём его фосфорилирования, либо удаления с поверхности клетки (эндоцитоз). Множество разных сигналов, воспринимаемых клеткой, суммируется в один определённый ответ.
Рецепторы по своей химической природе являются белками и, как правило, состоят из нескольких доменов. Рецепторы можно подразделить на:
|
МЕМБРАННЫЕ |
ВНУТРИКЛЕТОЧНЫЕ |
|
|
Механизм действия гормонов, взаимодействующих
с 7-ТМС-рецепторами
Наиболее распространенная группа рецепторов (рецепторы α- и β-адренергические, гистамина, серотонина, гликопротеиновых гормонов, глюкагона, паратирина, кальцитонина, гормонов гипоталамуса). Механизм усиления гормонального сигнала, реализуемого через данный вид рецепторов достигается синтезом небольших молекул - вторичных посредников (цАМФ, ИТФ, ДАГ, Са2+).
Рецептор состоит из 7 трансмембранных спиральных сегментов и соединяющих их гидрофильных вне- и внутриклеточных петлей. Последние содержат центры связывания с G-белками. Мембраносвязанный G-белок в неактивном состоянии — гетеротример (αβγ), α-субъединица которого связана с ГДФ.
Известно приблизительно 20 классов G-белков, но максимально распространены:
GS — активируют аденилатциклазу;
GI — ингибируют аденилатциклазу;
Gq — активируют фосфолипазу С.
Связывание гормона с рецептором изменяет конформацию последнего, что проводит к замене ГДФ в α-субъединице на ГТФ и последующей диссоциации G-белка (α + βγ). Передвигаясь по мембране α-субъединица взаимодействует с эффекторным белком-ферментом (аденилатциклазой, фосфолипазой С или каналом). Продолжительность эффекта определяется ГТФ-азной активностью G-субъединицы.
Gs-белки
α-субъединица Gs-белка контактирует и активирует мембранный фермент — аденилатциклазу, катализирующий образование из АТФ внутриклеточного проводника сигнала – цАМФ (рисю 20.1). Повышение его концентрации в клетке кратковременно, цАМФ под действием фосфодиэстераз распадается до 5'-АМФ.
цАМФ, в свою очередь активирует цитозольный фермент протеинкиназу А, гетеротетрамер из 2-х регуляторных и 2-х каталитических субъединиц. Присоединение цАМФ к регуляторным субъединицам вызывает высвобождение каталитических.
Каталитические субъединицы протеинкиназы А:
фосфорилируют ферменты, изменяя их каталитическую активность,
активируют, фосфорилируя, фактор транскрипции CREB, взаимодействующий с цАМФ-респонсивным элементом ДНК и усиливающий транскрипцию определенных генов,
вызывают открытие цАМФ-зависимых ионных каналов.
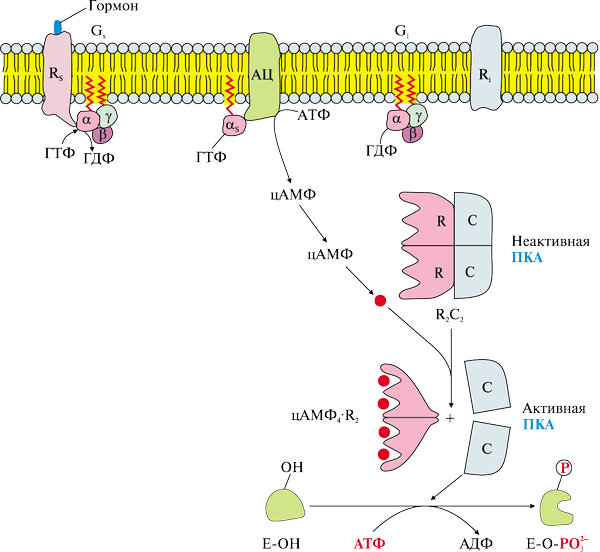
Рис. 20.1. Проведение гормонального сигнала через 7-ТМС-рецепторы
и аденилатциклазную эффекторную систему
Gq-белки
α-Субъединица Gq-белка контактирует и активирует мембранный фермент — фосфолипазу С, расщепляющую фосфатидилинозитолдифосфат на инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ).
ИТФ связывается со своим рецептором на кальциосомах (лиганд-зависимый кальциевый канал), что приводит к открытию канала и выходу в цитозоль ионов Са2+. Последние присоединяются к специальному связывающему белку кальмодулину или другим Са-связывающим белком и образующийся комплекс оказывает влияние на ряд ферментов (адеанилатциклазу, ФЛА2, множество киназ, NO-синтазу и др.), а также изменяет активность Са2+- насосов.
ДАГ, остающийся в мембране, либо активирует протеинкиназу С, фосфорилирующую и тем самым модулирующую активность различных ферментов, либо служит источником арахидоновой кислоты-субстрата для синтеза простагландинов.