

Osnovy_obschey_patologii_Zaychik_Churilov
.pdf! |
101 |
Рис. 6Связь. междунаследственност |
ью,влиянвнешнейсредыиреактивностьюем |
разаболеваниявитиипо(В.И.Арнольду). |
|
В — врачебноевоздействие; |
|
С — влияниевнешнейсреды; |
|
Р — реактивность; |
|
1, 2 — точкисостоянийсистемы: |
|
3,4 — траектоизмененияпарамеуправленияотк12ров. |
|
Этапроеимеетскладкициясборку.Совокупностьскладок |
— |
||
криваякатастроф.При |
[57] переходеэтойкривойчислопрообразов |
||
проекцийскачкообразноменяется.Нобудетли,зависитотк |
|
|
|
траекторииизменений, толькоотначальногоконечн го |
|
— определяетсянетолько |
|
пунктов. |
Тоесть,будетлиболезнь |
|
|
начальконечными з ачениемпарамет,которыеизменилисьов подвнешнвоздействием,нотемспособом,кот стигнуторым этоизменение.

! 102
Приизменениипараметровуправления( рис. 6)отточки1 точкепо2траекториипроизойдет3 пересечениекривойка,астроф ипроекцточкокажетнаикачеяновомсуровнеятвенно.
Этом уподобитьжнотакойкомбинацииС,ВРвпредыдущие моменты,когданаступилабол езньздоровый(организмс нормальнойиндивидуальнойреактипребольнойностьюратился, скачеосжизобойтвенедеятельностью).
Тотжепуоточкиь1точкемог2бытьпроделанподругой |
|
|
траекпри4,этолиниякатастрофриимнепереивсекается |
|
|
точкисохраняюттожечислопрообразов,тоестьпереходид т |
|
|
скачкаиболненаступаетзнь,хотязначенияСВпришликтому |
|
|
жеуровню,чтопервомварианте. |
|
|
Образного,зависимостиоряотреактивнос,индивидможети |
шестэтажанадцатого |
|
выбратьразличныепути,ведущиес |
|
|
первый:черезлифт,почернойлестнице…черезокно.Очевидно, |
|
|
чтод стоднихтехжегнувконечных, окажетсярдинат,в |
|
|
зависимостиотвыбранногопутистоимадаптации,весьмасти |
— здоров,состомя |
нии. |
разном,необязательно |
||
Всвететеориикатастрофреактивносвыглядиткак ь |
|
выбора системойтогоили |
индивидуальныйвнутренмеханизм |
|
|
иногопутивпространствеадаптивныхсостояний. |
|
|
СУБСТРАТРЕАКТИВНОСТИЭВОЛЮЦИЯ ЕЕИНТЕГРАТИВНЫХМЕХАНИЗМОВ
Хотяпонятиереактивность« »при |
|
надлежиторганизмукак |
||
цел,конкретмеханизмыу реактивностимогутыереализоваться, |
-листруктурнобоз |
-функциональных |
||
преимущественно,накаком |
||||
подуровней.Поэтомусубстратреактивностиможетбытьусловно |
|
|
емы.Говорят |
|
отнесенккакому |
-тоуровнюорганживойсистзации |
|
||
реактивностинамолекулярнклеточном,суб, ,тканевом иорганнуровне,выдрганизменныеляютдажепопуляционные формыреактивности.
Намолекулярномуровнереа тивностилючевоезначен е
! |
103 |
имеюткомплементарныевзаимодействия,ос |
нованные |
однозначномструктурномсоответствиираспознающихмолекул. |
|
Так,рецепторсубъединиаденилатциклазыаяспецифически узнаетпептидныйбиорегуляторвзаимодействуетнимпо принципуключ« -замок»,чтоведетак каталитическойивц субъединицыэтогофермента.Комплементарностьпроявляетсяво взаимодействферментовихсубстратов,антигеновантителях,
цис-регуэляторныхементовхроматиналигандов,изменяющих экспрессиюгенов.
[58] ВпослврработамиеднеемяДж. |
Блэлокаисоавторов |
|
||
(1986-1990)пока,чтовзакоиномноплементарныенитиДНК |
|
|||
кодируютбелковыемолекулы,котор,своюоче,преоявляютедь |
|
|
|
|
междусобойструктурноеоднозначноесоответствие.Иными |
|
|
|
|
слова,гориихмрецепторыионынапример( ,рецепторАКТГ |
уются,соответственно,смысловойи |
|
||
адренокортикотропин)кодир |
|
|||
антисмысловойРНК,которыесписанысимметричных |
|
|
|
|
комплементарныхкодирующучастковДНК.Еслэтотпринцип |
|
|
|
|
окажеуниве,тпсяорсальпризиде,чтраспознающиесяымать |
|
|||
молезакодирвулыгеномепоп«симметричварноны |
|
|
но»и |
|
каждбелкусоответствуетмукодегоантибелка |
-визавиилисвоего |
|
||
родааутоантитела« ». |
|
|
|
|
Вэтомслучаедревнепик,итайскуюостоящграммуиз ю |
но |
|||
взаикомноплементарныхфигурИнь«»Ян»« ( |
рис. 7)мож |
|||
будетсчитатьпророческимизображениемсмысловойструктуры |
|
|
|
|
геномаилипринципакомплементреактивности.Пок,чторнойзано |
возникактахтр ютнскрипцииЖ.( |
- |
||
антисмысловыеРНК |
||||
Ж. Тульме, и1986)служатрегулят,таккакспокрамисобны |
|
|
|
|
комплементарному блокированиютрансляциисмысловыхрепл к
{6}.

! |
104 |
Рис. 7Древнек. символИнь""Ян""тайский |
— символическоеизображение |
комплементарности |
|
Принципкомплементар,скоторыммыстолкуженостиулись прассмоти ренииэлемолекулярногоентарногоуровнясубстрата реактивности,носитсквознойхарактерпр насутствуетвсех урорганизациивняхживого.
Механизмыреактивности,свойственныееесубклеточному |
|
|
клеточнуровням,подробноохарактеризованымупоследующих |
|
|
разделах.Патология( сигнализации,Патологиярецепции.д.)В. |
|
|
данномобщемразделехотелосьбытолькопод,чтоеркнуть |
|
|
молекулярные,субклеточныеи механизмыреактивности |
|
|
ненасебеутпечатьиндивтойидуальностижемере,вкакойе |
|
|
высшие интегральныепроявления.Реактивностьраздтехичная |
|
|
илииныхмолекул,органоидовклеток,взяразличных |
|
|
индизразличныхвидовтканей.Фетальныйгемоглобин |
|
|
гемоглобинАпо |
-разномусвязываюткислор.Митохондрии |
качествеэнерг тических |
скелетныхмышцпредпочитаютв |
|
|
субстратактивныеоднофрагментуглероднполученные, из |
— фрагменты, |
|
глюкозы,амитохондриикардиомиоцитов |
||
полученТканевойизжирнкислот. ыкровянойхтромбопластин |
и |
|
различаютсяпосоставумеханизмамобразования.Макрофаг |
||
разныхтканей,напеченочные, римерклеткиКупф ра |
- |
|
остеокласты,несмотнаобщностьпроисхожя,выгляпо дятения |
||
разномуит.п. |
|
|
Следующимииерархическуровнямсубстратареактивностими
! |
105 |
являются тканевой и органный. |
|
Помередифференцировки,клеткиостав |
ляютвак ивно |
используепрограаппаратетолькомчастьмномгенетически унаследовпр, грстархивируютсяммльныенных.Поэтомуони отвечаютнараздрре ,свойсжениякциямиданнойтвека. ными Примеромпроявлтканевыхханизмовнияреактивностиможет служитьмесоваскуляризованныхветныйтканейнаповреждение
— воспаление.
Сразвитиеморганогенезаможносвязатьначалоформирования системныхответов,таккакаждыйоргансостоитизразличных тканей.
Важнойсостканевогоавляющейиорганногосубстрата реактивноявляетсяти структурно-функциональныйэлемент органа (ткани).
Несмотрянаразличияназванийнефрон(почек,печеночная |
— вмышцеит.д.) |
|
долька — впечени,двигательнаяединица |
||
структурно-функцедиоргановонитканцыимльныеютй |
М. Чернуху,микроциркуляторные |
|
общиечерты |
строения.ПоА. |
|
сосудистыеединицытипичногостроенияслужат |
[59] структурной |
|
осью,вокругкоторойгруппируютсясоединительно |
-тканные |
|
элементыстроморгана,вып полняющие,трофическуюную |
рганнойпаренхимы. |
|
защитнуюфункциюдляэлементов |
||
Структурно-функциэлементсоединительнойнальтканиый |
||
Чернухназвал« |
гистионом»При.любомповрежденииоргана |
|
специализировзащитфункциивовачаыеленныекаются |
|
|
элементыстроми,вполневозм,чтопринебольшихжно |
|
|
масштабахпроц |
ессэтимогра,невызываяичивается |
|
существенногорасстройствафункцийоргп ннойренхимы.Именно |
М. Чернуха,мобъяснитьжно |
|
этим,сточзренияконцепцииА. |
||
наличиебезжг патлтушных,клкоторыхиникатовпредставлена, |
ойзащитреакции.Знойачение |
|
основном,симптостромальнами |
||
этогооригинальногоподходамывидимто,чтогистионы |
- |
|
дейслужаттвительпервичаренойзащитной |
||
приспособреакц,такиепатойпроцессыьныхлогические,как |
— развертываются |
|
воспаление,гипер,иш, емия,тазромбоз |
||
! |
|
106 |
именновгистионах. |
|
|
Дублировадеятельномножеструктурноие ваи |
- |
|
функцед,составляющихницнальныхорган,определяет |
|
|
надежность системы,дажееслипригибелиэленементыогут |
|
|
регенерировать. |
|
|
Вздорганизмеовом,каку азываютАдоНовицкий, |
-15%паренхиматозных |
|
используется20 |
-25%нефронов, 12 |
|
элементовпечени.д. |
|
|
Всвязиэт,функциональныем пробы,оценивающие парциальфункциипочек,порнемыеовыявитьйгутнарушений больных,значительныйпроцентпочечнойпаренхимыкоторыхуже поражен ефросклерозом.
Науровнесистеморгановсоздаетсякачественноиная интегмехреактивностинцизмовя — вокругтехнологической задачитойилинойсистемы,приучастиивзаимодействий, описанныхП. К. Анохинымсм(.предыраз).дущийел
Восноведеятельности |
каждойизсистем,интегрирующих |
механизмыреак,будьтонервнаяивности,эндокриннаяили |
-таки,комплементарноевзаимодействие |
иммунная,лежит,опя ь |
|
регуляторасрецепторно |
-дискри.минаторнойстемЕгоскв зной |
характернеслучаен:однозначноесоответств |
иеодноймолекулы |
другойслужитформструктурноготраженияй.Аотражение |
— |
добиологическаяосновареактивности.Комплементарные |
|
взаимодействия,вер, ятзнираньшечемклетки.Покрайней |
— вирусы и |
мере,известныенамдоклеточформыжизниые |
|
прионы — способныккомплементарномувзаимодействию |
|
клетками.Прионыведутсебя,какнекиеомплементарные |
|
белковыесигналы,способныеприпопаданвклеткуактивировать |
|
древнейшиеубиквитарныегенетическиепрограммы, |
ов. |
обеспечивающиевоспроизводствоэтихагент |
|
Еслижизньначаласьприонов,чтовполневероятно,это означалобы,чток мплементарноевзаимодействиебылорешающим шагвеепомявлении.
! |
107 |
Понекоторымпредставозниклиниям,первымирецепторные белки,скореевсего,выполнявшиепоначалурольрепресс орных регуклеяпроцто.Необходимостьчныхровссовагента, снимающегоограничение,вызваладавлениеотборазакрепление мутаций,приводящихкобесродспечениюпротогормоновва рецепторов.Появилисьбелкби выеразнообразие, егуляторы которыхвдальн ейшемнарасталоприотносительномконсерватизме рецепторов.Возм,наобжноордестлястоероидныхло гормонов:тоестьразнообразрецепторовнарасталопри эволюционнойстабильностирегуляторов.
Иммуноглобулины,возникшиенаосновегомологичных |
|
|
распознающихбелков,представленныхуорганизмов,не |
|
|
обладающихиммуннойсистем,оказалисьвсостоянииобеспечитьй |
|
|
уникальноеразнообразиерегуляторныхсигн.Какужелов |
|
|
отмечалосьвыше,реактивностьможетрассматриватьсякак |
). |
|
дискнаборретныйактонов( |
функциональныхфенов |
|
Функционкаждтакедобеспечиваетийрованницыинд виду |
|
|
преивсмывоущеспроизводствале, жевремяимеет |
|
|
определецену.Прималыхиннуютенсивностяхиспользованиятого |
го,нопри |
|
илииногореактонавыигрышрастетбысплзнеарееты |
|
|
высоких — ростплатыопережаетприросвы,этогрышаделает |
|
|
присспомданнсореактоблениещьюпасгов( нпоаымним |
|
|
определеГеккелюболезпо !)нипатогеннымподробнее( см. |
|
|
А. Н. Горбань,Р. |
Г. Хлебопрос, 1988). |
|
Нореактивностьвключае тнетолькореаи, ктоны функциусткомбийчивыенальсоздаваемыереактонов, ации интеграаппаратомпо( ерминологииивнымК. Люкаса(1909) — включаетспособифункциональноеповедениеости,использующее ихпутемкомбинации).
Новыереактоныэволюции возникаютнамногореже,чемновые функциональныеомбинацстарыхмозаблоковчных (А. М. Уголев, Р.1985)Гийменназвалэтооппортунизмом.
эволюционпроцессаилиприиспн«ципомогот,чтользуйд рукой» (1984).
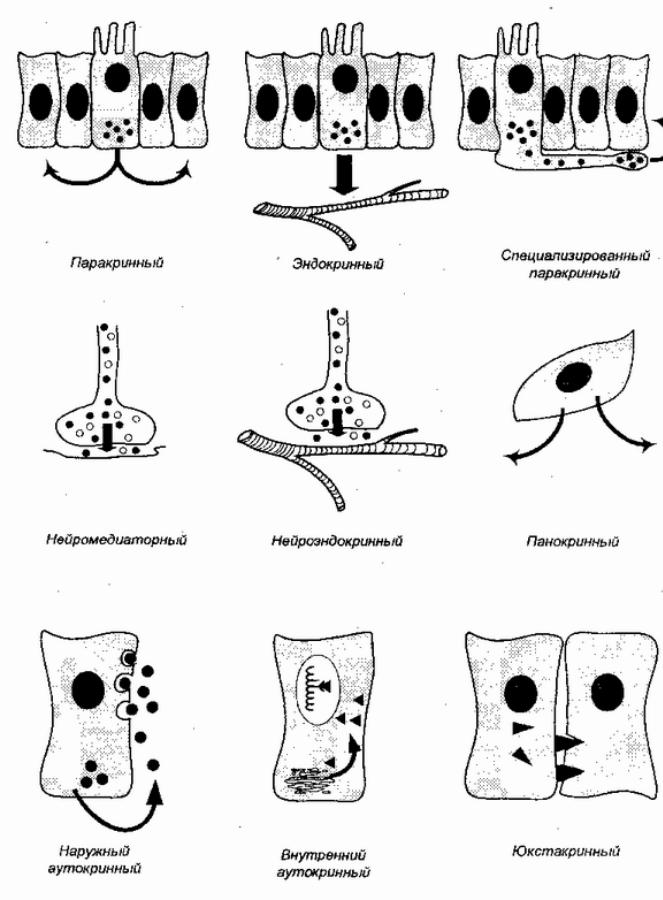
! |
108 |
Рис. 8Различные. типыхимическихре |
гуляторныхвзаимеждуклод йствийтками |
|
(поГ. |
Дж. ДокреюиК. |
Р. Хопкинсу, 1982) |
! |
|
|
|
|
109 |
|
|
|
Тиротропгормонобнаруженбактерий,ыйинсулинего |
|
— упростейших,а |
||||||
рецептор — удрозофил,тропныегормоны |
|
|||||||
нейропептиды — угубок,неимеющихЦНС.Морфинизмакаимее |
|
|
т |
|||||
кодируемыесходгеаналогиымиамивдиффузнойэндокринной |
|
|
|
|||||
системеживотных |
|
— эндорфэнкефалины, орфины |
Трабукки, 1978)Это . |
|
||||
(Дж. Хьюз, X. |
Костерлиц, 1975,Э. |
Коста,М. |
Снайдер, |
|||||
жеотникопсирецепторамтсяатнымЛ.( |
|
Терениус,С. |
||||||
Э. Саймон, 197 |
4). |
|
|
|
||||
Вазопрессинумлекопидейснаобитающихвуетр,ательныеубки |
|
Ю. Йейтс, 1982). |
||||||
ауамфибий |
|
|
|
— наклеткикожФ. ( |
|
|||
Биохимическиеэлементырегулясисэвтемлюционируютрных |
|
|
|
|||||
какцелоеВ(. |
|
|
Ле Руа, 1987). |
|
|
ьнового |
||
Следовательно,эволюцияреактивностииспользуетпут |
|
|
||||||
поведенческогоиинтеграткомбстарыхивногонирования |
эволюционныеизобретения |
|
||||||
эволюционныхнаходок,реактоныка |
|
|
||||||
непропадают,хотяивостребуютсяпоройдляиныхфункций. |
|
|
|
|||||
Эволюциявыступаеткакмножественныевариациинаогранич нное |
|
|
|
|||||
числотем |
|
-архетипов. |
|
|
|
|||
Ф. Ницшеоснэтомовалпринципесвойизвесафоризм,такный |
|
|
|
|||||
раздражающийнекоторыхистоп ическихогрессистов |
|
|
|
|||||
оптимиЛюди(«,проделавстпучервяйткьчел, овеку |
|
|
|
|||||
многомосталисьчервями»). |
|
|
|
|
регуляции |
|||
Филогдревнейшийнетспособхимическойчески |
|
|
||||||
клеточнойреактивности |
— аутокринный.Онможетбыть |
|
||||||
внутре,когданнимоклеточныйорганизмвырабатывает |
|
|
|
|||||
химическиерегуляторы,действующие,нвыходяизклетки.Есл |
|
|
|
|||||
биорегувыдедейлянасторетсяамтввырабатывающуюетего |
|
|
|
|||||
клетку,говорят |
|
|
|
онаружнойтокриннойрегуляции.Аутокринные |
|
|
||
воздействияинтерлейкина |
-2 — важныйэлементкооперацииклеток |
|
||||||
виммунномответе.Когдабиорсегуляторыкретируютсявовне |
|
|
|
|||||
действуютнадругиеклетки,проявляетсямеханокринныйизм |
Р. |
Хопкинсу, При1982). |
|
|||||
(см. |
рис. |
8,поГ. |
Дж. ДокреюиК. |
|
||||
|
|
|
|
|
|
|
|
|
панокриннойрегуляцклеткавыделяб иорегулятор окружающеепространство,жидкуюсредуоб литания. чву Такспсуществуетойсободноклеточныхипростых многоклеточных,напримерлесневыегрибыводоросливыделяют антиби,подавляярбактерийсттики.Ж антибиотиквотный лизоцим,присутствующийпрактическивосехо етахганизма,
! |
|
|
110 |
|
|
можетрасцениваться,какнаследиепанокриннойрегуляции |
|
|
|
||
реактиворганизости |
|
ма.Юкстаквоздействиеприннедп, лагает |
- |
||
чтолипофильныйбиорегулятороставмембранеклеткится |
|
||||
источникаактивируеконтакклеткойсяом |
|
|
-мишенью. |
||
(М. Списоавт.рн1992). |
|
|
|
|
|
Следующимэволюционнымшагоммоглобытьвозникновение |
Фейртера, регуляции1938) |
|
|
||
паракринной (терминФ. |
|
|
|||
многоклеточных.Припаракринномвоздействиибиорегулятор |
|
|
|
||
выделяетсяобластитесныхклеточныхконтактовивлияетлишь |
|
|
|
||
нареакцииближайшихсоседейклетки,связанныхнеюэтими |
|
|
древняя |
||
цитологическструктура.Ворганчеловекаизмеи |
|
|
|||
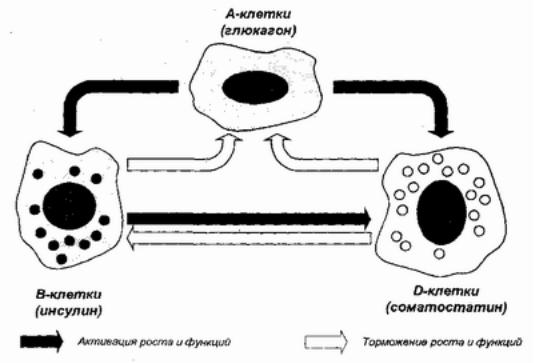
паракриннрегуляциясохр.Примероманяетсямогслужить |
|
|
|
||
взаимоотношенияосновныхклеточныхэлементовостровка |
|
|
— |
||
Лангергансавподжелезеелудочной.Триосновныхтипаклеток |
|
|
|||
α,β,иδобразуютгетери,помимоклеточныевыделения |
|
|
— в |
||
своихбиорегу |
ляторов — глюкагона,инсулинасом тостатина |
|
|||
системныйкровоток,качествегормонов,испоихдльзуютя |
|
|
|
||
локальвозднруейсдругаочерезотвияесныеклеточные |
[62] |
|
|
||
конткакп ракриннктысигналы. |
е |
|
матостатин |
||
Какпаракрклеточныеыеси,иг иалысулин |
-клеток( |
|
|||
ингибсекрециюп уютолиферациюα |
рис. 9). |
||||
|
|
|
|
|
|
Рис. 9Паракринное. взаимклетокстровкадействиеЛангерпо(Унгарунса |
-Орчи) |