

Глава 24
МОТОРИКА И СЕКРЕЦИЯ В ПИЩЕВАРИТЕЛЬНОМ ТРАКТЕ
L
ПЕРЕДВИЖЕНИЕ ХИМУСА
Передвижение пищевого комка (а в дальнейшем — химуса) по желудочно-кишечному тракту — процесс, который осуществляется на оральном и аборальном концах с участием произвольных поперечно-полосатых мышц, а на остальных этапах — с участием гладкой мускулатуры. Заглатывание пищи и акт глотания требуют обязательного участия централь ной нервной системы, поэтому выделяют центры жевания, центры глотания. Жевание осу ществляется жевательными мышцами (иннервация тройничным нервом), мимическими мышцами (лицевой нерв — VII пара), а также мышцами языка (XII пара, подъязычный нерв). Эфферентные нейроны этих черепномозговых нервов вместе с нейронами ретикулярной формации, красного ядра, черной субстанции и коры больших полушарий составляют так называемый центр жевания. Именно он обеспечивает координированное действие мышц — участников акта жевания. Регуляция силы, необходимой для развития усилий при откусы вании, раздавливании, растирании пищи, осуществляется с участием проприорецепторов этих мышц. г
Акт глотания — достаточно сложный процесс, цель которого — перевести пищевой комок в пищевод и одновременно перекрыть все пути для попадания пищевого комка в носовые пути, в дыхательные пути или назад — в ротовую полость. В акте глотания выделяют 3 фазы: 1) ротовая произвольная, которая заключается в том, что комок пищи перемещается на корень языка, где находятся механорецепторы, вызывающие развитие следующей фазы; 2) глоточная, быстрая непроизвольная фаза, которая существует для того, чтобы перекрыть пути для пищевого комка в дыхательные и носовые пути; 3) пищеводная, медленная непроизвольная — в результате этой фазы открывается глоточнсьпищеводный сфинктер, и пищевой комок поступает в начальные области пищевода.
Перемещение пищевого комка на корень языка — это один из произвольных элементов акта жевания, благодаря мышцам языка и мимической мускулатуре пищевой комок переносится на корень языка. В глоточной фазе участвуют различные структуры: рецепторы ротовой полости, в том числе механорецепторы корня языка. Информация от них идет по афферентным волокнам тройничного нерва, языкоглоточного и вагуса (гортанный нерв) и возбуждает эфферентные нейроны вагуса, языкоглоточного и подъязычного нервов — нейронов, которые вместе с нейронами среднего мозга и коры больших полушарий составляют основу центра глотания. Итак, поток эфферентных импульсов вызывает следующие три процесса: 1) закрытие входа в носовую полость за счет приподнимания мягкого неба и язычка; 2) закрытие входа в гортань — для чего происходит поднятие гортани и смещение ее вперед путем смещения подъязычной кости; 3) закрытие входа в ротовую полость — поднятием вверх корня языка.
3-я фаза заключается в том, что в момент акта глотания пищевод подтягивается к зеву. Начальная его часть расширяется и принимает пищевой комок. С этого момента начинается этап перемещения пищевого комка по пищеводу. Осуществляется это при помощи так называемой перистальтики. Такой тип движения широко распространен в ЖКТ — он обусловлен тем, что в сторону желудка бежит волна сокращения циркулярной мускулатуры пищевода. Как у желудка, тонкого и толстого кишечника, так и у пищевода имеются два основных гладкомышечных слоя — продольный (наружный) и циркулярный (внутренний).
329
Каждый из слоев может сокращаться самостоятельно, независимо от другого слоя. При перистальтике в основном сокращается циркулярная мускулатура, а продольная лишь способствует этому процессу. Обычно первичная волна перистальтики доходит до места пересечения пищевода с аортой, здесь зарождается вторичная волна сокращения, которая передвигает пищевой комок к кардии. В области кардии находится нижний пищеводный сфинктер (кардиальный сфинктер), который при приближении пищевого комка расслабляется и позволяет войти ему в желудок.
Важную роль в обеспечении односторонности движения и предотвращении заброса пищи из желудка в пищевод имеет острый угол впадения пищевода в желудок (так называемый угол Гиса). При наполнении желудка его острота возрастает, что препятствует регургита-ции содержимого желудка в пищевод. Примерно такую же роль играет и газовый пузырь в дне желудка — он тоже уменьшает угол Гиса. Кроме того, переходу химуса из желудка в пищевод препятствуют косые циркулярные мышцы желудка и диафрагмально-пищеводная связка (мембрана Лаймера).
В среднем твердая пища проходит пищевод за 8—9 с, жидкая за 1—2 с. Организация волны сокращения (перистальтики) осуществляется, вероятно, за счет межмышечного, или ауэрбахова сплетения (по А. Д. Ноздрачеву — это метасимпатическая нервная система). Волокна вагуса при их активации усиливают перистальтику пищевода и одновременно расслабляют тонус кардии желудка. Симпатические волокна, иннервирующие пищевод, при возбуждении оказывают противоположный эффект. Гуморальные механизмы, по-видимому, здесь не играют какой-либо значительной роли.
ЖЕЛУДОК: ДЕПОНИРОВАНИЕ И ЭВАКУАЦИЯ ПИЩИ
Во время акта глотания происходит расслабление гладкой мускулатуры желудка — это явление называется пищевой рецептивной релаксацией. Она существует на протяжении почти всего промежутка приема пищи и обусловлена, вероятнее всего, влиянием вагуса на метасимпатическую гастральную систему. Как будет показано ниже, вагус как парасимпатический нерв обычно вызывает повышение активности — за счет деполяризации гладких мышц желудка и кишечника. Каким же образом в этот период вагус вызывает расслабление гладких мышц? В специальной литературе этот вопрос обходят молчанием. Можно предположить, что либо есть нехолинергические постганглионарные волокна вагуса (в момент акта глотания возбуждаются некоторые нейроны вагуса, которые передают сигнал на нехолинергические постганглионарные нейроны), либо эффект вагуса, а точнее нейронов, обслуживающих акт глотания, опосредуется через эфферентные нехолинергические нейроны ауэрбахова сплетения (через метасимпатическую нервную систему). Идея о существовании различных популяций нейронов вагуса, участвующих в регуляции моторики желудка и, возможно, кишечника, например, нейронов, обеспечивающих релаксацию во время глота-ния или поддержания тонуса мышц во время пищеварения, достаточно хорошо объясняет тот факт, что моторика кишечника и желудка бывает различной по своему характеру и функциональному назначению, а вагус всегда способствует соответствующей активности.
Итак, после пищевой рецептивной релаксации наступает повышение тонуса, необходимого для того, чтобы пища начинала перетираться, сжиматься, смешиваться с желудочным соком. Одновременно моторика желудка должна обеспечить процесс перемешивания химуса, перевода слоев, лежащих внутри химуса, в соприкосновение со стенкой желудка и наоборот. Этот процесс осуществляется за счет перистальтических волн, которые примерно 3 раза в минуту возникают в области кардии и со скоростью 1 см/с распространяются в сторону пилорического отдела. В начале процесса пищеварения такие волны слабые, редкие, но по мере окончания пищеварения, в желудке (что, очевидно, оценивается хемо-, ме-хано- и осморецепторами желудка) они возрастают как по интенсивности, так и по частоте. В результате такой перистальтической волны небольшая порция химуса подгоняется к ан-труму желудка для того, чтобы выбросить ее в последующем из желудка. По своей природе
330
эта перистальтическая волна обусловлена преимущественно сокращением циркулярной мускулатуры желудка, волна этих сокращений всегда направляется в одном и том же направлении — к антруму. Следовательно, она каким-то образом организована. Можно утверждать, что организатором такого движения является метасимпатическая система (ауэрбахово сплетение) за счет местных периферических рефлексов. Это доказывается, в частности, тем, что ваготомия, которая не так уж редко проводится у людей с целью, например, лечения язвенной болезни желудка, не сопровождается нарушением такого механизма. Вероятно, вагус может только регулировать вид активности этого сплетения (выше указывалось, что при глотании степень сокращений уменьшается, наступает релаксация). Известно, что характер перистальтики зависит от вида пищи, консистенции, объема. Все эти факторы могут «разумно» отразиться на характере перистальтики лишь в том случае, если информация от механо-, хемо- и осморецепторов будет передаваться на местные рефлекторные дуги.
Регуляция перистальтики осуществляется и гуморальными факторами, продукция которых во многом зависит от вида и объема пищи. Усиливают перистальтику гастрин, моти-лин, серотинин, инсулин, а тормозят ее — секретин, холецистокинин-панкреозимин, глю-кагон, ЖИП, ВИП, бульбогастрон.
По мере освобождения 12-перстной кишки и переваривания пищи периодически в желудке возникает особая перистальтическая волна, которая достигает такого повышения внутрижелудочного давления, при котором открывается пилорический сфинктер и порция химуса переходит в 12-перстную кишку. Такую волну называют перистальтической волной пропульсивного характера, или пропульсивной волной, или систолическим сокращением пилоруса. Волна эта возникает в районе кардии, но наибольшее сокращение наблюдается в области пилоруса. Благодаря этой волне давление в области пилоруса возрастает до 20—30 см водного столба и становится выше давления в 12-перстной1сишке. В реализации этой волны участвуют циркулярные мышцы и, частично, продольные. Сокращаясь, продольная мускулатура как бы уменьшает фундально-пилорический размер желудка, что вместе с циркулярной перистальтической волной создает более высокое давление, чем при обычной перистальтике. В специальной литературе не описаны механизмы, лежащие в основе организации этого вида моторной активности желудка. Можно предположить, что организует этот процесс особый тип осцилляторов метасимпатической нервной системы, для возбуждения которых необходима суммация афферентных импульсов от различных рецепторов желудка и 12-перстной кишки. Ситуация напоминает активацию инспираторных нейронов типа la-для того, чтобы совершился вдох, необходим поток импульсов от хеморецепторов (меду-лярных и периферических). Представление о существовании особого осциллятора основа-, но на известных фактах: переход химуса из желудка в 12-перстную кишку зависит от объема, состава, консистенции, степени измельченности и разжиженности, осмотического давления, температуры, рН, состояния химуса в 12-перстной кишке. Состояние в 12-перстной кишке — очень важный фактор: закисление среды, накопление или даже появление в этой области жира (его сложно переваривать) — все это тормозит эвакуацию пищи. По мнению Г. Ф. Коротько (1987), регуляция перехода химуса осуществляется за счет рефлексов, которые замыкаются на уровне ЦНС, на уровне экстра- и интрамуральных ганглиев, а также модулируется интестинальными гормонами.
Когда у человека удаляется привратник и соответствующий сфинктер, то эвакуация пищи все равно осуществляется порционно. Это лишний раз говорит о том, что имеется осциллятор, предназначенный для организации процесса эвакуации. Он находится, вероятнее всего, в области кардии, откуда в норме начинаются пропульсивные волны.
Рвота представляет рефлекторный непроизвольный процесс, характеризующийся выбросом химуса из желудка в ротовую полость. Рвота возникает при раздражении ряда рефлексогенных зон: рецепторов корня языка, глотки, слизистой желудка, кишечника, брюшины, а также при чрезмерном раздражении вестибулярного аппарата. На основе сочетаний индифферентного раздражителя с безусловным сигналом (возбуждением указанных рефлексогенных зон) возможна выработка условнорефлекторной рвоты.
331
По механизму рвота представляет собой следующий процесс: сокращается тонкая киш* ка, т. е. возникает антиперистальтическая волна, и она движется от тонкого кишечника до пищевода. В результате химус забрасывается в желудок, затем кардиальный сфинктер рас-* слабляется. Одновременно возникают сильные сокращения мышц брюшной стенки и дна' фрагмы — на выдохе содержимое выбрасывается. При этом закрывается вход в носовую полость, дыхательные пути.
Акт рвоты — это сложно организованный процесс, в который вовлечены различные структуры: гладкие мышцы тонкого кишечника, желудка, пищевода, поперечно-полосатые мышцы живота, диафрагмы, носоглотки. Центр рвоты находится на дне IV желудочка, в области скопления ядер DC, X, XII пар нервов и связан с мотонейронами мышц живота и диафрагмы, с нейронами дыхательного центра и метасимпатической нервной системой кишечника и желудка. Почему возникает антиперистальтическая волна? Очевидно, что в процессе эволюции была выработана реакция, позволяющая избегать недоброкачественную пишу, и сформировались механизмы, обеспечивающие антиперистальтическую волну. Вероятнее всего, это метасимпатическая система (ауэрбахово сплетение), которая включается при возбуждении осциллятора, расположенного в области тонкого кишечника. Возбуждение этого осциллятора запускается нейронами вагуса, ответственными за акт рвоты. На этом примере видно, что в желудочно-кишечном тракте есть набор «стандартных программ» с единым осциллятором. Каждый такой набор, например, «акт рвоты», «акт эвакуации химуса из желудка в 12-перстную кишку» запускается в действие с помощью специализированных нейронов вагуса (нейроны рвоты, нейроны эвакуации и т. п.).
Не случайно А. Д. Ноздрачев вводит понятие о модулях в энтерииовой метасимпатической системе: набор стандартных программ реализуется за счет комплекса модулей, имеющихся в желудочно-кишечном тракте. Очевидно, что в эволюции потребность в существовании таких модулей возникает в том случае, когда деятельность регулируемой системы многообразна и управлять ею из единого центра сложно. Сочетание централизма (решение общих или «системных» задач) и регионарных задач — вот, вероятно, удачная идея природы. Все сказанное здесь можно распространить и в отношении регуляции процессов секреции, в которой принимает участие мейсснерово сплетение.
МОТОРИКА ТОНКОГО КИШЕЧНИКА
Она представлена несколькими видами двигательной активности: 1) ритмическая сегментация, 2) маятникообразные сокращения, 3) перистальтические сокращения (очень медленные, медленные, быстрые, стремительные), 4) антиперистальтические сокращения, 5) тонические сокращения. В тонком кишечнике совершается перемешивание химуса, что улучшает процесс переваривания пищи и постепенное передвижение химуса в аборальном направлении (к толстому кишечнику). Это передвижение может быть различным по скорости (очень медленным, медленным, быстрым, стремительным), что, вероятно, определяется уровнем возбудимости нейронов, обеспечивающих реализацию модуля «перистальтики». Все типы движений: маятникообразные движения, перистальтика и антиперистальтике — обусловлены особым образом организованными нейронными сетями ауэрбахова сплетения с единым водителем или водителями ритма, которые в зависимости от ситуации возбуждены или заторможены. Характер моторной активности тонкого кишечника во многом определяется видом пищи, интенсивностью гидролиза и всасывания. Комплекс местных рецепторов: механо-, хемо-, температурных, осморецепторов — обеспечивает соответствующие потоки афферентных импульсов, которые и вызывают активацию в данной области кишки соответствующего модуля. Вагус как представитель центра определяет принципы функционирования и регулирует степень активности соответствующего модуля.
Ритмическая сегментация — это периодическое сокращение циркулярных мышц кишки. Благодрая этому кишка делится на отдельные сегменты, т. е. возникают перетяжки. Это вызывает перемешивание химуса.
332

Рис. 90. Кимограмма моторики кишечника лягушки и влияния на нее ацетилхолина. пилокарпина и адреналина:
I— исходная двигательная активность кишечника;
— влияние ацетилхолина;
— влияние поликарпина;
IV — влияние адреналина: а — кривые сокраще ния; 6 — отметка времени с ценой деления 5 с. Стрелками отмечены моменты нанесения рас творов на кишку. Ацетилхолин использовался в разведении 1.1000, пилокарпин —1:100, адрена лин—1:100.

Рис. 91. Кимограмма изолированного отрезка кишечника кролика. Влияние на нее раздражения блуждающего и симпатического нервов:
I — при раздражении блуждающего нерва,
II — при раздражении симпатического нерва; а — запись моторики, б — отметка времени с ценой деления 30 с.
Стрелками отмечены моменты нанесения раздражения.
Маятннкообразные сокращения —
сочетанное сокращение циркулярных и продольных мышц, в результате чего химус перемещается «вперед-назад» и одновременно с малой скоростью движется в аборальном направлении. Этот вид движения способствует перемешиванию химуса.
Перистальтические движения возникают в результате распространения волны сокращения циркулярной мускулатуры в аборальном направлении при одновременном расслаблении продольной мускулатуры, расположенной ниже цир-кулярно активного участка. Благодаря этому виду движения химус перемещается в аборальном направлении со скоростью 0,1—3 см/с. Скорость перистальтического движения определяется видом пиши, а также состоянием тонуса ЦНС — вагус усиливает, симпатические нервы тормозят.
ГГ. Г. Богач в 1961 г. сформулировал закон моторной деятельности пищеварительного тракта: адекватный раздражитель вызывает возбуждение моторики в данном месте и ниже этого места, а активность вышерасположенных участков при этом тормозится. Поэтому существуют такие рефлексы, как пищеводно-ки-шечный (при прохождении пищи по пищеводу происходит активация моторики кишечника, в том числе и толстого), желудочно-кишечный (возбуждается активность кишечника и тормозится активность мышц пищевода), ректоэнтераль-ный (при возбуждении толстого кишечника активность тонкого кишечника тормозится). По мнению П.Г. Богача, дуги этих рефлексов замыкаются в ЦНС, симпатических брюшных ганглиях и интра-муральных ганглиях.
Моторная активность тонкого кишечника повышается под влиянием многих факторов: кислоты, щелочи, концентрированные растворы солей, продукты гидролиза, серотинин, гистамин, гаст-рин, мотилин, холецистокинин-панкре-озимин, вещество Р, вазопрессин, бра-дикинин, препараты, повышающие хо-линомиметическую активность вагуса, например, прозерин.
333
(О моторике желчного пузыря — см. Желчевыделение.) МОТОРИКА ТОЛСТОГО КИШЕЧНИКА
Переход химуса из подвздошной кишки в слепую происходит через илеоцекальный сфинктер (баугиниева заслонка). Он устроен по типу клапана — пропускает химус лишь в одном направлении. Вне пшцеварения сфинктер закрыт. После приема пищи каждые 1—4 минуты он на некоторое время открывается (в среднем на 0,5—1,0 минуту) и химус (по 15 мл) поступает в слепую кишку. Когда давление в слепой кишке превышает давление в подвздошной, сфинктер не открывается, и поэтому химус из слепой кишки не поступает в тонкий кишечник. Это важно и для микрофлоры — ее перемещение из толстого кишечника в тонкий не происходит.
Анализ моторики толстого кишечника показывает, что если рентгеноконтрастную массу (бариевую кашицу) ввести в желудок, то в толстом кишечнике она обнаруживается уже через 3—3,5 часа, то есть весь пассаж химуса — бариевой кашицы совершается за 3—3,5 часа. В последующие 24 часа происходит заполнение толстого кишечника, а полное опорожнение происходит за 48—72 часа.
Таким образом, основное время, которое затрачивает химус на продвижение по желудочно-кишечному тракту, — это время на пассаж через толстый кишечник, в котором происходит окончательное всасывание веществ, переваривание неразрушенных питательных веществ с участием микроорганизмов и формирование каловых масс.
Для толстого кишечника характерны следующие виды движения:
1) маятникообразные (малые и большие), 2) перистальтические, 3) антиперистальтические и 4) пропульсивные, т. е. перистальтические движения большой интенсивности, благодаря которым происходит заполнение каловыми массами прямой кишки. Пропульсивные движения возникают до 3—4 раз в сутки и могут вызывать позывы на дефекацию.
Принцип организации двигательной активности толстого кишечника такой же, как и для желудка и тонкого кишечника: метасимпатическая система, организованная в виде отдель ных модулей: нейроны-осцилляторы, задающие ритм для соответствующего модуля; влия ния из центра в виде импульсов от вагуса (активация) или от симпатических волокон (угне тение активности). ■&
Позыв на дефекацию возникает при повышении давления в прямой кишке до 40—50 см водного столба, а при давлении 20—30 см водного столба возникает чувство наполнения прямой кишки. Акт дефекации представляет собой сочетание следующих процессов: 1) расслабление внутреннего (непроизвольного) анального сфинктера, 2) расслабление наружного (произвольного) анального сфинктера, 3) возникновение перистальтики прямой кишки пропульсивного типа, 4) повышение внутрибрюшного давления с участиеммышц брюшной стенки и диафрагмы за счет натуживания до 220 см водного столба. Регуляция этих процессов осуществляется с участием нейронов, локализованных в пояснично-крестцовом отделе спинного мозга (центр непроизвольной дефекации), а также с участием нейронов продолговатого мозга, гипоталамуса, коры больших полушарий (центр произвольной дефекации), которые оказывают свой эффект через центр непроизвольной дефекации. Суть регуляции сводится к тому, что в ответ на поток импульсов от механорецепторов (рецепторы растяжения) прямой кишки возрастает тонус нейронов парасимпатической нервной системы в области крестцового отдела спинного мозга — это расслабляет непроизвольный сфинктер и усиливает моторику кишечника. Одновременно возбуждаются альфа-мотонейроны поясничного отдела, которые вызывают расслабление произвольного сфинктера и активируют мышцы, участвующие в натуживании.
В желудочно-кишечном тракте известно примерно 35 сфинктеров, или клапанов, которые играют исключительно важную роль в процессах пассажа пищи и химуса по ЖКТ. Нарушение работы этих сфинктеров порой вызывает такие существенные изменения в функции ЖКТ, что больные погибают или находятся в тяжелом состоянии. Правильная трактов-
334
ка роли сфинктерного механизма позволила курганскому хирургу профессору Я. Д. Витебскому предложить много новых оперативных вмешательств на желудке и кишечнике по созданию анастомозов между отдельными областями ЖКТ с учетом физиологии клапанного аппарата ЖКТ.
Согласно Г.Ф. Коротько (1987), различают верхний сфинктер пищевода, нижний (карди-альный) сфинктер пищевода, пилорический сфинктер желудка, бульбодуоденальный сфинктер, сфинктер Хелли добавочного протока поджелудочной железы, сфинктер Одди-Бойде-на общего желчного протока, сфинктер Вестфаля главного протока поджелудочной железы, сфинктер Шройбера большого дуоденального сосочка, сфинктер пузырного протока Люткенса, сфинктер печеночного протока Мириззи, сфинктер Капенджи, сфинктер Оксне-ра (оба — в области тонкого кишечника), дуоденальная связка Трейца, сфинктер илеоце-кальный Варолиуса (илеоцекальный клапан), сфинктер червеобразного отростка, или заслонка Герхала, сфинктер Бузи (в области слепой кишки), сфинктер Гирша (начало восходящей ободочной кишки), сфинктер Кеннона (поперечно-ободочная кишка), сфинктер Хо-рета (средняя ее часть), сфинктер Кенюпа—Беша (перед спуском), сфинктер Пайра—Штрауса (начальная часть нисходящей толстой кишки), сфинктер Блалли—Михайлова (в конце нисходящей части толстой кишки), сфинктер Росс—Мютье (сигмовидная кишка), сфинктер О'Берна—Пирогова—Мютье (начало ректум), третий ректальный сфинктер, внутренний сфинктер ректум, наружный сфинктер ректум.
СЕКРЕТОРНАЯ ФУНКЦИЯ ЖКТ
Секреция различных соков, слюны — важнейшая функция ЖКТ. Слюнные железы, пан* креатическая железа — это обособленные железы, в которых вырабатываются секреты. Эти секреты идут в полость ЖКТ. Существует множество железистых клеток, которые находятся в толще слизистой ротовой полости, желудка, тонкого и толстого кишечника, в которых осуществляется секреция, продукты которой выделяются в полость ЖКТ через специальные мелкие выводные протоки. Это мелкие слюнные железы, желудочные железы, брун-неровы железы 12-перстной кишки, либеркюновые железы тонкой кишки, бокаловидные клетки тонкого и толстого кишечника. Отдельное место занимает печень: ее гепатоциты, выполняя одновременно ряд других процессов (обезвреживание, метаболизирование продуктов), продуцируют желчь, которая необходима для переваривания жиров как активатор и эмульгатор.
Секреция в ЖКТ обеспечивает наличие в секретах гидролитических ферментов, создание оптимальной рН среды, наличие защитных факторов (слизь, бактерицидные вещества). Секреторные клетки ЖКТ выполняют и роль экскреторных структур. По происхождению секреты являются продуктами анаболизма (собственно секрет) и катаболизма клетки (экскрет) и продуктами, которые поглощаются клеткой, а затем ею же выделяются, т. е. являются проходящими «транзитом» через секреторную1 клетку (рекрет).
Процессы секреции протекают в три основные фазы:
1) поступление исходного материала — воды, аминокислот, моносахаридов, жирных кислот, что осуществляется путем пассивного и активного транспорта этих частиц из кро-вотока; 2) синтез первичного секреторного продукта и его транспорт для секреции. Согласно Г. Ф. Коротько (1987), в панкреатических клетках во 2-ю фазу из поступивших в клетку аминокислот на рибосомах эндоплазматического ретикулюма в течение 3—5 минут происходит синтез белка-фермента (в неактивной форме). Затем этот белок в составе пузырьков переносится в аппарат Гольджи (на это затрачивается около 7—17 минут), где он пакуется в вакуоли, в которых гранулы профермента транспортируются до апикальной части секреторной клетки, здесь совершается 3-я фаза — выделение секрета, или экструзия (экзоци-тоз). От начала синтеза до выхода секрета проходит в среднем 40—90 минут.
В ЖКТ характер выделения секрета мерокриновый, т. е. клетка не разрушается, а выделяет упакованно секрет через специальные отверстия в апикальной мембране. Секрет может также покинуть клетку за счет диффузии.
335
Все три фазы секреции регулируются, т. е. можно изменить поступление исходных продуктов, скорость синтеза, объем и скорость выделения секрета. Это осуществляется при помощи гуморальных влияний, в основном, интестинальнымн гормонами, а также за счет нервных влияний со стороны местных рефлекторных дуг, локализованных в мейсснеровом (подслизистом) сплетении, т. е. за счет метасимпатической нервной системы и влияний со стороны ЦНС, которые реализуются через вагус и симпатические волокна. На все воздействия секреторная клетка, которая относится к возбудимым тканям, отвечает изменением уровня мембранного потенциала. Факторы, усиливающие секрецию, обычно вызывают деполяризацию клетки, а тормозящие секрецию — гиперполяризацию. Деполяризация обусловлена повышением натриевой и снижением калиевой проницаемости мембраны секреторной клетки, а гиперполяризация обусловлена повышением хлорной или калиевой проницаемости. Мембранный потенциал у клетки вне периода секреции составляет SO мВ, причем МП апикальной и базальной мембран разный, что имеет значение для направленности диффузионных потоков. В момент секреции возникает временная деполяризация клетки — секреторный потенциал.
Описанный механизм характерен, главным образом, для действия медиаторов симпатической, парасимпатической и метасимпатической нервной системы. Для гормонов более характерной является их способность изменять интенсивность секреции и выделение секрета за счет посредников — цАМФ или цГМФ (гормон + рецептор -> активация протеинки-назы ->...-> конечный эффект) или за счет изменения внутриклеточной концентрации кальция (кальций + кальмодулин -> активация протеинкиназы ->...-> конечный эффект).
Все виды регуляции базируются на информации, идущей от рецепторов пищеварительного канала. Механо-, хемо-, температурные и осморецепторы по афферентным волокнам вагуса, языкоглоточного нерва, а также по местным рефлекторным дугам дают информацию в ЦНС и метасимпатическую нервную систему об объеме, консистенции, степени наполнения, давлении, рН, осмотическом давлении, температуре, концентрации промежуточных и конечных продуктов гидролиза питательных веществ, концентрации некоторых ферментов.
Регуляция секреции осуществляется за счет прямого влияния на секретируемые клетки и опосредованно, например, за счет изменения кровотока продукции местных интестиналь-
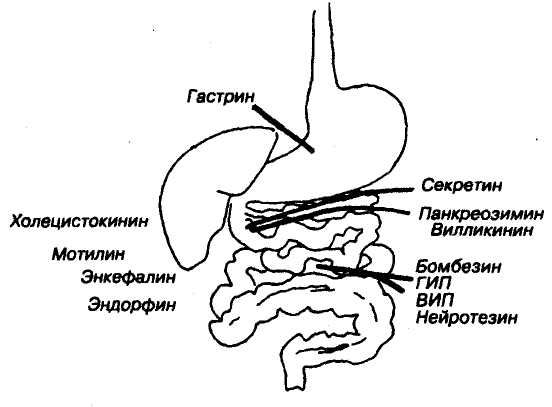
Рис. 92. Роль гуморальных факторов в регуляции деятельности пищеварительного тракта.
ных гормонов, активности нейронов метасимпатической системы, регулирующих, в свою очередь, активность секреторных клеток. Центральные механизмы регуляции секрецин — это нейроны коры больших полушарий (существует множество условных пищевых рефлексов, в том числе секреторных), нейроны лимбической системы, ретикулярной формации, гипоталамуса (передние и задние ядра), продолговатого мозга. В продолговатом мозгу среди парасимпатических нейронов вагуса имеется скопление нейронов, которые непосредственно реагируют на
336


Рис. 93. Рефлекторная регуляция желудочного сокоотделения.
А — афферентные нервы, эфферентные парасимпатические и симпатические нервы. Пм — продолговатый мозг, Гл—симпатический ганглий, См—спинной мозг, Ж—желудок, Яз—язык. Б — схема условного рефлекса на желудочное сокоотделение. Условный раздражитель — свет, подкрепление — пища. Выработанный условный рефлекс: действие света приводит к выделению желудочного сока К—кора, П -г подкорковые центры, Зц—центры зрения, Пц— пищевые центры, Ж — желудок.
а фферентные и эфферентные (от коры, гипоталамуса и пр.) потоки импульсов и посылают эфферентные сигналы к симпатическим нейронам, расположенным в спинном мозге, и одновременно — к секреторным клеткам ЖКТ. Возможно, что часть волокон вагуса непосредственно взаимодействует с секреторными клетками, а часть (основная) — опосредованно, через взаимодействие с эфферентными нейронами метасимпатической нервной системы. Существование возможности выработки условных секреторных рефлексов на соответствующие виды пищевых продуктов позволяет предполагать, что отдельные популяции вагуса регулируют активность соответствующего модуля метасимпатической нервной си-