

15.8. Функциональная специализация мозговых структур в образовании эмоций
При осуществлении каждой из четырёх функций эмоций решающая роль принадлежит одной из четырёх структур мозга:
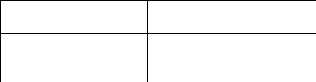
Поведенческие функции эмоций Структуры, сохранность которых
имеет критическое значение для
данной функции
Отражательно- оценочная Фронтальная и височная области коры
Компенсаторная (замещающая) Гиппокамп
Переключающая Миндалины
Подкрепляющая Гипоталамус
С помощью позитронно-эмиссионной томографии было установлено, что мысленное представление эмоционально нейтральных событий сопровождается преимущественной активацией дорсолатеральной префронтальной коры и задней височной области, тогда как во время мысленного представления эмоциональных переживаний активируются орбитофронтальная кора и передняя височная область. Предполагают, что лобная доля правого полушария преимущественно связана с ранее приобретённой информацией, необходимой для удовлетворения потребности, а левая лобная доля - с информацией, поступившей только что, при этом рассматриваются не все, а лишь наиболее вероятные сигналы. При сравнении этих двух видов информации возникает эмоциональная оценка события.
Гиппокамп тоже относится к информационным структурам, его задача связана с извлечением следов памяти о прежнем опыте, на основании которого оценивается вероятность удовлетворения потребности. Мотивационное возбуждение гиппокампа осуществляет гипоталамус, часть сигналов поступает также от перегородки. При нарастании эмоционального напряжения в условиях дефицита информации гиппокамп увеличивает количество извлекаемых из памяти энграмм, тем самым стимулируя поисковое поведение. Благодаря этому реакция на любой сигнал, высоковероятный или маловероятный, становится почти такой же, как, например, на сигнал явной опасности (т.е. по типу доминанты), а поэтому ни одно действительно важное сообщение не пропускается. Такое замещающее недостаток информации реагирование сопровождается выраженными вегетативными изменениями ("вегетативная буря"), вызванными симпатоадреналовой реакцией.
При конкуренции между существующими мотивациями возникает необходимость выбора доминирующей потребности, которая удовлетворяется в первую очередь. Этот выбор осуществляют миндалины, он зависит от внешних условий в конкретной ситуации и в конкретный момент времени. На основании позитронно-эмиссионной томографии можно предполагать, что свою переключающую функцию миндалины реализуют при участии хвостатых ядер.
Гипоталамаус является важнейшей мотивационной структурой, но одновременно он осуществляет подкрепляющую функцию эмоций. Эта функция выражается в стремлении к эмоционально положительным стимулам и избеганию эмоционально отрицательных, т.е. к максимизации или минимизации соответствующих эмоций.
Многолетние исследования механизмов возникновения эмоций и формирования поведения, выполненные П. В. Симоновым и его коллегами позволили обнаружить индивидуальные особенности относительной активности информационных (фронтальная кора и гиппокамп) и мотивационных (миндалины и гипоталамус) структур. В результате было высказано предположение о том, что индивидуальные отношения между информационными и мотивационными структурами определяют такой параметр поведения, как экстраверсия-интроверсия (Рис.15.6). Одновременно с этим соотношение систем фронтальная кора - гипоталамус и гиппокамп - миндалина определяет, возможно, индивидуальную особенность поведения, которую принято рассматривать как эмоциональная стабильность - нейротизм.