

14.3. Кибернетические принципы гомеостатического регулирования
Биологические потребности связаны с сохранением важнейших гомеостатических параметров: если они по какой-либо причине отклонятся от заданного значения, то автоматически срабатывают нервные и эндокринные механизмы регуляции и одновременно изменяется поведение: оно мотивируется на такие действия, которые помогают быстрее восстановить гомеостаз. Регулирование гомеостатических параметров (например, парциального давления кислорода в артериальной крови, осмотического давления крови, уровня сахара в крови, величины артериального давления и т.д.) можно рассмотреть с позиций теории управления - одного из разделов кибернетики, которая изучает информационные процессы в сложных системах. Системой в кибернетике называется совокупность взаимодействующих между собой относительно элементарных структур или процессов, объединённых в целое для выполнения общей функции. Например в биологических системах, управление необходимо для поддержания оптимального значения того или иного гомеостатического параметра. Если повлияют внешние силы и изменят величину такого параметра, то входящие в систему звенья перестраиваются так, чтобы вернуть ему оптимальное значение (Рис. 14.2).
Рис. 14.2. Общая схема гомеостатического регулирования
Запускающие деятельность системы внешние стимулы
+/- +/-
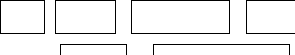 Заданное
Управляющее
Вегетативная регуляция
Реальное
Заданное
Управляющее
Вегетативная регуляция
Реальное
значение устройство Эндокринная регуляция значение
параметра +/- (мозг) Поведение параметра
+/-
Обратная связь Измерение величины параметра
Если, например, заданное значение средней температуры тела близко к 37, а внешняя температура (запускающий стимул) начинает повышаться (+) или понижаться (-), то соответствующая информация от наружных терморецепторов заставляет центры температурной регуляции (управляющее устройство) воздействовать на механизмы нервной и эндокринной регуляции, а также на поведение так, чтобы они и в этих условиях помогли сохранить имеющуюся температуру. Если же это не удаётся, то температура тела начнёт увеличиваться или уменьшаться, т. е. заданное значение параметра изменится. Величину отклонения от заданного значения определят внутренние терморецепторы (измерение величины параметра) и оповестят об этом (обратная связь) центры температурной регуляции. После этого активность центров меняется так, чтобы выровнять регулируемый параметр по заданному эталону. По такой же схеме кроме температуры тела регулируются и многие другие гомеостатические параметры, отклонение которых от заданного значения автоматически запускает внутренние механизмы регуляции (1) и в то же время специфически изменяет характер поведения (2).
14.4. Гипоталамус - важнейшая мотивационная структура мозга
В кибернетическом понимании гипоталамус является управляющим устройством, к которому поступает вся необходимая информация о сохранении гомеостатических параметров или их отклонении от заданного значения. На языке физиологических терминов это означает получение афферентных сенсорных сигналов от периферических рецепторов, а также способность центральных рецепторов самих гипоталамических нейронов непосредственно воспринимать изменения состава крови и ликвора.
Эти два источника поступающей в гипоталамус информации стали основой двух физиологических теорий мотиваций: периферической и центральной. В основу периферической теории легли исследования, показывавшие закономерную связь между мотивированным поведением и характером импульсации от определённых периферических рецепторов: сухость во рту, например, связывалась с формированием жажды, а периодические сокращения пустого желудка - с возникновением голода. В середине ХХ века ведущую роль в формировании мотиваций начали отдавать гипоталамическим центрам, которые в то время открывали один за другим. Позже оба подхода естественным путём объединились, поскольку информация от периферических рецепторов попадает, как известно, в гипоталамические центры и специфически изменяет их активность.
В середине 50-х годов существовала точка зрения, что роль гипоталамуса в формировании мотиваций сводится лишь к общему сенсорному и моторному возбуждению, но вскоре стали находить специфические мотивационные механизмы голода и насыщения, питьевого и температурного поведения и т.д. Эти находки были получены во время наблюдений за последствиями электрической стимуляции отдельных регионов или ядер гипоталамуса, а также за изменениями поведения экспериментальных животных после электролитического разрушения тех или иных областей с помощью введённых в гипоталамус электродов.
В гипоталамусе имеется обильная сеть кровеносных капилляров, ни одна другая область мозга так густо ими не насыщена. Через стенки этих капилляров способны проходить такие растворённые в крови вещества, которые в других областях мозга никогда не попадут в его ткань из крови (или наоборот) в связи с особенностями строения стенок сосудов и расположением клеток глии, формирующими гематоэнцефалический барьер: в области гипоталамуса этот барьер снижен. Нейроны гипоталамуса имеют специфические рецепторы для связывания некоторых компонентов крови. Так, например, у одних клеток гипоталамуса существуют глюкорецепторы, взаимодействующие с молекулами глюкозы, а у других - рецепторы, связывающие тот или иной гормон.
Академик К. В. Судаков на основе многолетнего опыта исследования механизмов биологических мотиваций сформулировал представление о том, что различные группы гипоталамических нейронов используют для своих обменных процессов только определённые гуморальные факторы, т.е. обладают химической избирательностью. Если их внутренняя потребность в таком веществе не удовлетворяется, они приходят в состоянии возбуждения. Различные группы однородных в своей химической избирательности нейронов образуют разные мотивационные центры гипоталамуса. Специализация нейронов может состоять и в том, что одни из них активируются снижением уровня глюкозы в крови, а другие - повышением, есть нейроны, чувствительные к величине осмотического давления, уровня норадреналина в крови и т.д.
Возбуждение нейронов мотивационных центров происходит постепенно: по мере нарастания метаболической потребности достигается критический уровень их деполяризации и вследствие этого нейроны начинают ритмически генерировать потенциалы действия до тех пор, пока потребность не будет удовлетворена (Рис. 14.3). Это триггерный механизм деятельности, которая происходит периодично: нейроны ритмически разряжаются при возникновении потребности и успокаиваются, когда потребность будет удовлетворена.
Мотивационные центры гипоталамуса взаимодействуют с ретикулярной формацией, активность которой повышает общий уровень бодрствования и этим способствует эффективности действий, направленных на удовлетворение потребности. Гипоталамус имеет двусторонние связи со структурами лимбической системы мозга - именно этим определяется эмоциональная окраска мотивированного поведения. Наконец, у гипоталамуса существуют двусторонние связи с корой больших полушарий, в особенности с лобными долями, необходимыми как для создания двигательных программ, так и для определения эмоциональных аспектов поведения . Если возникает метаболическая потребность, то больше других структур чувствительный к этому гипоталамус активирует все связанные с ним области мозга, выполняя тем самым функцию пейсмекера - водителя ритма их активности. В свою очередь, связанные с гипоталамусом структуры мозга способны тормозить или усиливать его активность. Особая роль в этом отношении принадлежит коре, тормозящие влияния которой могут подавить или отсрочить удовлетворение той или иной потребности: например, многие верующие люди сознательно ограничивают себя в еде во время поста.
Изучение влияния нейротрансмиттеров на нейроны гипоталамуса показало, что разные вещества по-своему изменяют характер поведения. Так, например, нанесение норадреналина на паравентрикулярные ядра гипоталамуса сильно стимулирует аппетит, причём, при возможности выбирать ту или иную пищу, подопытные животные предпочитают продукты с высоким содержанием углеводов. Точно так же действует и самый сильный возбудитель аппетита - нейропептид Y, а ещё один пептид - галанин избирательно повышает потребление жиров.
Некоторые вещества, способные изменять характер поведения, попадают в гипоталамус из крови, проходя через гематоэнцефалический барьер. Так, например, ангиотензин, образующийся в повышенном количестве при уменьшении объёма циркулирующей крови, проникает в гипоталамус и активирует в нём определённые нейроны, что приводит в конечном счёте к усиленному потреблению воды. Некоторые пептидные гормоны образуются в двенадцатиперстной кишке и верхнем отделе тонкого кишечника в связи с поступлением туда пищи (например, холецистокинин, бомбезин и т.п.). Они попадают в кровь в очень небольших количествах, однако определённые нейроны гипоталамуса к этому высоко чувствительны: действие таких гормонов ведёт к уменьшению или прекращению приёма пищи. К таким же последствиям приводит и действие гормона поджелудочной железы инсулина, способного медленно проникать из крови в ликвор: в гипоталамусе найдены места связывания инсулина.
Таким образом, гипоталамус, обладающий всей информацией о состоянии внутренней среды организма, способен управлять её гомеостатическими параметрами с помощью нервных и эндокринных механизмов регуляции на основе уже имеющихся, накопленных ранее резервов. Но, кроме того, гипоталамус ещё и побуждает другие области мозга специфически изменить поведение, направить организм на такие действия, которые обеспечат восстановление потраченных ресурсов или их заготовку впрок.