

Внутриклеточное переваривание различных химических соединений и структур происходит в лизосомах. Лизосомы – это небольшие овальные тельца диаметром около 0,4 мкм, окружённые одной трёхслойной мембраной. В лизосомах находится около 30 ферментов, способных расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и другие вещества. Лизосомы могут разрушать и структуры самой клетки при её отмирании в ходе эмбрионального развития, когда происходит замена зародышевых тканей на постоянные, и в ряде других случаев. По-видимому, переваривание структур, образованных самой клеткой, играет важную роль в нормальном обмене веществ клеток. Однако остаётся неизвестным, каким образом лизосомы «распознают» внутриклеточный материал, подлежащий разрушению.
В различных метаболических циклах участвуют ферменты, содержащиеся в пероксисомах, состоящих из пузырьков диаметром 0,1–1,5 мкм, ограниченных одной мембраной. Пероксисомы ликвидируют возникающие в клетке пероксиды, токсичные для живого вещества.
Включениями называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
9.4.2. Жизненный цикл клетки
В многоклеточном организме клетки специализированы, т. е. имеют строго определённое строение и функции. В соответствии со специализацией
клетки имеют разную продолжительность жизни. Например, нервные и мышечные клетки после завершения эмбрионального периода развития перестают делиться и функционируют на протяжении всей жизни организма. Другие клетки, такие как клетки костного мозга, эпидермиса, эпителия тонкого кишечника в процессе своей специфической функции быстро погибают, и поэтому в этих тканях происходит непрерывное клеточное размножение.
Закономерные изменения структурнофункциональных характеристик клетки во времени составляют содержание жизненного цикла клетки (клеточного цикла). Клеточный цикл – это период существования клетки от момента её образования путём деления материнской клетки до собственного деления или гибели. Важным компонентом клеточного цикла является митотический цикл – комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткой многоклеточного организма специфических функций, а также периоды покоя. В периоды покоя ближайшая судьба клетки не определена: она может либо начать подготовку к митозу, либо приступить к специализации в определённом функциональном направлении (рис. 9.2). Митоз – способ деления клеток, обеспечивающий тождественное распределение генетического материала между дочерними клетками и преемственность хромосом в ряду клеточных поколений.
Из рисунка видно, что после завершения митоза клетка может вступить в период подготовки к синтезу ДНК,
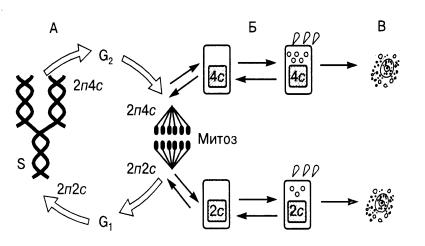
обозначаемый символом G1. В течение этого периода в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в биосинтезе ДНК. После завершения фазы G1 клетка приступает к синтезу ДНК или её редупликации – удвоению. Продолжительность синтеза ДНК в разных клетках неодинакова: от нескольких минут у бактерий до 6–12 часов в клетках млекопитающих.
Рис. 9.2 Жизненный цикл клетки многоклеточного организма: А – митотический цикл; Б – переход в дифференцированное состояние; В – гибель клетки (2п – двойной набор хромосом; 2с – количество ДНК в двойном наборе хромо-
сом; 4с – удвоенное количество ДНК).
После завершения синтеза ДНК – S-фазы митотического цикла – клетка, как правило, начинает делиться не сразу. Период от окончания синтеза ДНК и до момента митоза называется фазой G2. В этот период завершается подготовка клетки к митозу. Для осуществления митотического
деления клетки необходимы и другие подготовительные процессы, в том числе удвоение центриолей, синтез белков, из которых строится ахроматиновое веретено, завершение роста клетки. При вступлении клетки в митоз меняется её функциональная активность: например, прекращается амёбоидальное движение у простейших и у лейкоцитов высших животных; поглощение жидкости и деятельность сократительных вакуолей у амёб; часто исчезают специфические структуры клетки, например, реснички эпителиальных клеток.
Митоз (рис. 9.3) состоит из четырёх фаз: профазы, метафазы, анафазы и телофазы. В профазе увеличивается объём ядра, хромосомы становятся видимыми вследствие спирализации, по две центриоли расходятся к полюсам клетки, между ними микротрубочки образуют веретено деления. Вследствие спирализации хромосом становится невозможным считывание генетической информации с ДНК и прекращается синтез РНК. В конце профазы ядерная оболочка распадается на отдельные фрагменты, края которых смыкаются. Образуются мелкие пузырьки, сходные с эндоплазматической сетью. На протяжении профазы продолжается спирализация хромосом, которые становятся толстыми и короткими. После распада ядерной оболочки хромосомы свободно и беспорядочно лежат в цитоплазме.
В метафазе спирализация хромосом становится максимальной и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Митотическое веретено деления уже полностью сформировано и его микротрубочки соединяют полюса с центромерами хромосом. Каждая хромосома продольно
расщепляется на две хроматиды (дочерние хромосомы), соединённые в области центромеры. Центромерные участки хромосом находятся строго в экваториальной плоскости, а дочерние центромеры и хроматиды обращены к противоположным полюсам.
Рис. 9.3. Схема митоза. А, Б –интерфаза; В, Г – профаза;
Д– метафаза; Е – анафаза; Ж, З – телофаза:
1– центромера; 2 – ядрышко; 3 – центриоль; 4 – хромосома; 5 – ядерная оболочка; 6 – веретено.
Ванафазе центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными
хромосомами. Трубочки веретена, прикреплённые к центромерам, тянут хромосомы к полюсам клетки со скоростью 0,2–5 мкм/мин. По завершении движения на полюсах собирается два равноценных полных набора хромосом.
Завершается митоз телофазой. Хромосомы, собравшиеся у полюсов, деспирализуются и становятся плохо видимыми. Реконструируются ядра дочерних клеток. Образуются ядрышки. Из мембранных структур цитоплазмы образуются ядерные оболочки. Материнская клетка делится на две дочерние.
Биологическое значение митоза огромно. Благодаря точному распределению генетического материала между дочерними клетками митоз обеспечивает такие важные явления жизнедеятельности, как эмбриональное развитие, рост, восстановление органов и тканей после повреждения, поддержание структурной целостности тканей при постоянной утрате клеток в процессе их функционирования.
9.4.3.Структурно-функциональная организация генетического материала
Генетический материал, содержащийся в клетке, образует структурно дифференцированные единицы, называемые хромосомами. Хромосомы представляют собой мультимолекулярные агрегаты, образованные преимущественно молекулами ДНК и белка и содержащие небольшое количество РНК, не являющейся, строго говоря, структурной частью хромосомы.
Строение хромосом хорошо видно на стадии метафазы
митоза. Изучение хромосом позволило установить следующие факты: 1) во всех соматических клетках любого растительного или животного организма число хромосом одинаково; 2) в половых клетках содержится всегда вдвое меньше хромосом, чем в соматических клетках данного вида организмов; 3) у всех организмов, относящихся к одному виду, число хромосом в клетках одинаково, например, у человека в соматических клетках имеется 23 пары хромосом, а у голубя – 40. Число хромосом в соматических клетках всегда чётное, так как в них находятся по две одинаковых по форме и размерам хромосомы: одна от отцовского организма, а другая – от материнского. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, носит название
двойного, или диплоидного, набора. В половые клетки из каждой пары хромосом попадает только одна, поэтому хромосомный набор в этом случае называется одинарным
или гаплоидным.
Схематично строение хромосом представлено на рис. 9.4.
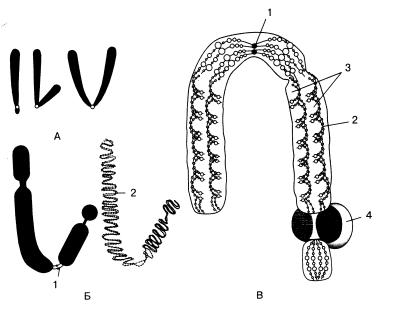
Рис. 9.4. Строение хромосом. А – типы хромосом; Б, В – тонкое строение хромосом:
1 – центромера; 2 – спирально закрученная нить ДНК; 3 – хроматиды;
4– ядрышко.
Вопределении формы хромосом большое значение имеет положение так называемой первичной перетяжки или центромеры – области, к которой во время митоза прикрепляются трубочки веретена. Центромера делит хромосому на два плеча. Расположение центромеры определяет три основных типа хромосом: 1) равноплечие –
сплечами равной или почти равной длины; 2) неравноплечие, имеющие плечи неравной длины; 3) палочковидные – с одним длинным и вторым очень коротким, иногда с трудом обнаруживаемым плечом.
Непосредственным носителем наследственной информации в хромосомах является ДНК
(дезоксирибонуклеиновая кислота) – биологический полимер, состоящий из двух полинуклеотидных цепей, соединённых друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырёх азотистых оснований: аденин (А), или тимин (Т), или цитозин (Ц), или гуанин (Г); сахар – дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов (рис. 9.5).
Азотистое |
|
|
|
|
Остаток |
||||
основание |
|
|
|
фосфорной |
|||||
аденин, |
|
Дезокси- |
|
|
кислоты |
||||
|
|
|
|
|
|
|
OH |
||
|
|
|
|
|
|
|
|||
|
рибоза |
|
|
|
|
|
|||
или гуанин, |
|
|
|
P |
|
|
О |
||
|
|
|
|
||||||
|
|
|
|
|
|||||
или цитозин, |
|
|
|
|
|
|
|
|
OH |
или тимин |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 9.5. Схема строения нуклеотида
В каждой цепи нуклеотиды соединяются путём образования ковалентных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. Объединяются две цепи в одну молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящих в состав нуклеотидов, образующих разные цепи. Количество таких связей между разными азотистыми основаниями неодинаково, и вследствие этого они могут соединяться только попарно: азотистое основание А одной цепи полинуклеотидов всегда
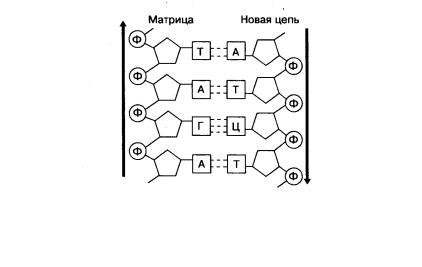
связано двумя водородными связями с Т другой цепи, а Г – тремя водородными связями с азотистым основанием Ц противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов называется комплементарностью. Комплементарное взаимодействие нуклеотидов приводит к образованию пар нуклеотидов (рис. 9.6). В полинуклеотидной цепочке соседние нуклеотиды связаны между собой через сахар (дезоксирибозу) и остаток фосфорной кислоты. В 1953 году американским биофизиком Дж.Уотсоном (род. 1928) совместно с английским биофизиком и генетиком Ф.Криком (род. 1916) была предложена модель пространственной структуры ДНК в виде двойной спирали
(рис. 9.7).
Рис. 9.6. Комплементарное соединение нуклеотидов и образование двухцепочечной молекулы ДНК:
А– аденин; Г – гуанин; Т – тимин; Ц – цитозин; Д – дезоксирибоза;
Ф– остаток фосфорной кислоты
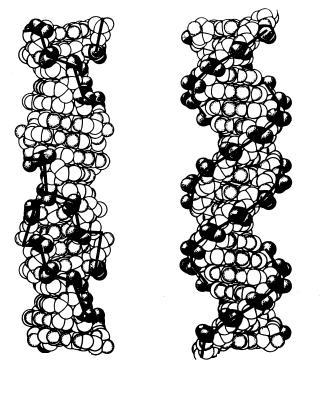
а |
б |
Рис. 9.7. Пространственные модели левозакрученной (а) и правозакрученной (б) спиралей ДНК.
Таким образом, в структурной организации молекулы ДНК можно выделить первичную структуру –
полинуклеотидную цепь, вторичную структуру – две
комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединённые водородными связями, и третичную структуру – трёхмерную спираль. Диаметр спирали составляет 2 нм, длина шага – 3,4 нм. В каждый виток входит 10 пар нуклеотидов. Длина спирали молекула ДНК зависит от организма, которому она принадлежит.
ДНК простейших вирусов содержит несколько тысяч нуклеотидных пар, бактерий – несколько миллионов, а высших организмов – миллиарды. Если выстроить в одну линию все молекулы ДНК, заключённые в одной клетке человека, то получится нить длиной 2 м, т. е. её длина в миллиард раз больше её толщины.
Относительная простота структуры ДНК, представляющей чередование всего лишь четырёх различных нуклеотидов, долгое время мешала исследователям рассматривать это соединение как материальный субстрат наследственности и изменчивости, в котором должна быть зашифрована чрезвычайно разнообразная информация. В 1954 году Г.А.Гамовым было высказано предположение, что кодирование генетической информации в молекулах ДНК должно осуществляться сочетанием нескольких нуклеотидов.
Всё многообразие жизни обусловливается разнообразием белковых молекул. Только в организме человека встречается около 5 млн типов белковых молекул, отличающихся по своей массе, структуре и функциям не только друг от друга, но и от белков других организмов. Однако, несмотря на такое разнообразие и сложность строения, все белки построены всего из 20 различных аминокислот. Для шифровки такого их числа достаточное количество сочетаний нуклеотидов обеспечивает
триплетный код, в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами. В этом случае из четырёх нуклеотидов образуется 43 = 64 триплета. Код, состоящий из двух нуклеотидов, дал бы возможность зашифровать только 42 = 16 различных аминокислот. Из 64 возможных триплетов ДНК кодирует аминокислоты 61 триплет, а оставшиеся 3 триплета выполняют функцию знаков препинания при считывании наследственной информации.
Генетический код обладает специфичностью. Каждый триплет способен кодировать только одну определённую аминокислоту. Интересным фактом является полное соответствие кода у различных видов живых организмов. Такая универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции.
Одним из основных свойств материала наследственности является его способность к самокопированию – редупликации. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе редупликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной спирали ДНК образуются две идентичные двойные спирали дочерних ДНК (рис. 9.8).
В процессе синтеза ДНК принимает участие целая группа ферментов. С помощью фермента геликазы разрываются водородные связи и двойная спираль ДНК расплетается в местах начала редупликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые