

Межвидовая конкуренция в природе
В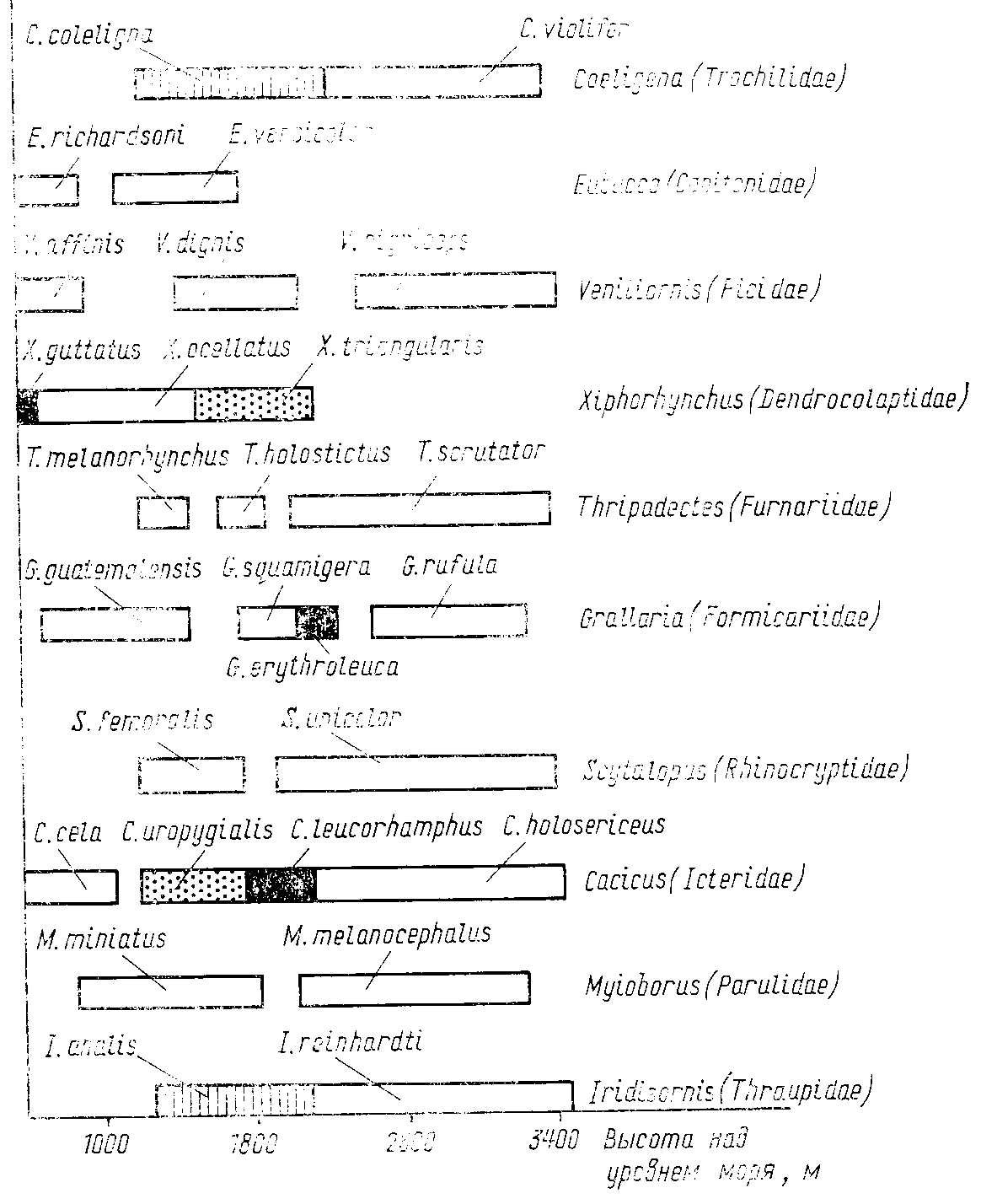 последние два десятилетия в экологической
литературе ведутся ожесточенные споры
по поводу того, какова роль конкуренции
в ограничении распространения и динамики
природных популяций разных видов, а,
следовательно, и в определении структуры
сообществ. По мнению одних исследователей,
популяции, входящие в естественные
сообщества, достаточно строго
контролируются системой конкурентных
отношений, иногда, правда, модифицируемых
воздействием хищников. Другие же
полагают, что конкуренция между
представителями разных видов наблюдается
в природе только эпизодически, а
популяции, в большинстве своем, будучи
ограничены другими факторами, как
правило, не достигают тех плотностей,
при которых конкурентные отношения
становятся определяющими. Существует
также и не лишенная оснований компромиссная
точка зрения, предполагающая наличие
некоторого континуума реальных природных
сообществ, на одном конце которого —
сообщества, стабильные во времени,
богатые, или, точнее, насыщенные видами,
жестко контролируемые биотическими
взаимодействиями, а на другом—сообщества
нестабильные (в большинстве случаев
из-за того, что не стабильны абиотические
условия в их местообитаниях), не насыщенные
видами (т, е. допускающие вселение новых
видов) и контролируемые, как правило,
плохо предсказуемыми изменениями
внешних условий.
последние два десятилетия в экологической
литературе ведутся ожесточенные споры
по поводу того, какова роль конкуренции
в ограничении распространения и динамики
природных популяций разных видов, а,
следовательно, и в определении структуры
сообществ. По мнению одних исследователей,
популяции, входящие в естественные
сообщества, достаточно строго
контролируются системой конкурентных
отношений, иногда, правда, модифицируемых
воздействием хищников. Другие же
полагают, что конкуренция между
представителями разных видов наблюдается
в природе только эпизодически, а
популяции, в большинстве своем, будучи
ограничены другими факторами, как
правило, не достигают тех плотностей,
при которых конкурентные отношения
становятся определяющими. Существует
также и не лишенная оснований компромиссная
точка зрения, предполагающая наличие
некоторого континуума реальных природных
сообществ, на одном конце которого —
сообщества, стабильные во времени,
богатые, или, точнее, насыщенные видами,
жестко контролируемые биотическими
взаимодействиями, а на другом—сообщества
нестабильные (в большинстве случаев
из-за того, что не стабильны абиотические
условия в их местообитаниях), не насыщенные
видами (т, е. допускающие вселение новых
видов) и контролируемые, как правило,
плохо предсказуемыми изменениями
внешних условий.
П
Рис. 57. Смена видов одного рода в
зависимости от высоты над уровнем моря,
прослеженная в Андах для представителен
10 семейств птиц (по Terborgh, 1971)
Исследуя изменения видового состава птиц в Перуанских Андах по мере подъема в горы, Я. Терборгх (Terborgh, 1971) обнаружил, что виды одного рода очень четко сменяют друг друга, причем границы распространения часто не связаны с вертикальной поясностью растительности, а определяются, вероятно, только конкуренцией между близкими видами. На схеме (рис. 57), заимствованной из работы Я. Терборгха видно, что чем больше видов одного рода встречено на всем обследуемом диапазоне высот, тем меньший интервал высот приходится в среднем на один вид. Так, если от высоты 1000 до высоты 3400 м встречаются два представителя одного рода, то на каждого приходится интервал в 1200 м, а если в таком же диапазоне высот обитают три вида одного рода, то на каждый вид приходится в среднем по 800 м. Подобное распределение явно указывает на конкуренцию, и оно вряд ли может быть объяснено без учета межвидовых взаимодействий (MacArthur, 1972). Важные дополнительные свидетельства наличия конкуренции в случае, описанном Я. Терборгхом, были получены при изучении вертикального распределения птиц, проведенном при участии того же автора (Terborgh, Weske, 1975) в Андах, но не на основном хребте, а на небольшом изолированном горном массиве, расположенном от него в 100 км. Число обитающих здесь видов было значительно меньше, чем на хребте, но те же самые виды встречались в большем диапазоне высот, указывая на то, что именно конкурентные отношения, а не абиотические факторы ограничивают их распространение на основном хребте.
Много примеров межвидовой конкуренции дает островная фауна (Майр, 1968), представители которой нередко демонстрируют взаимоисключающее распределение, хотя на материке живут бок о бок. Так, М. Радованович (Radovanovic, 1959; цит. по: Майр, 1968), изучив распространение ящериц рода Lacerta на 46 островах Средиземного моря около побережья Югославии, выяснил, что на 28 островах встречалась только Lacerta melisellensis, а на остальных — только Lacerta sicula. Нет ни одного острова, на котором оба вида обитали бы вместе.
В более редких случаях исследователи могли непосредственно наблюдать расширение области распространения одного вида, сопровождающееся исчезновением или сокращением численности в этой области другого вида, являющегося его потенциальным конкурентом. Так, с конца XIX вплоть до середины XX в. в Европе было замечено резкое сокращение ареала широкопалого рака (Astacus astacus) и соответствующее расширение на северо-запад ареала близкого вида — длиннопалого рака (Astacus lepiodactylus), захватившего весь Волжский бассейн, а затем проникшего в бассейн Невы и Северского Донца (Бирштейн, Виноградов, 1934). В настоящее время оба вида встречаются в Прибалтике и Белоруссии, однако, случаи их нахождения в одном водоеме очень редки (Цукерзис, 1970). Механизм вытеснения одного вида другим не ясен, за исключением тех немногих случаев, когда длиннопалый рак был специально запущен в те водоемы, где широкопалый погиб при эпизоотии «рачьей чумы» — грибкового заболевания, способного полностью уничтожить популяцию речных раков. Вполне вероятно, что успешному расширению ареала A. lepiodactylus способствовало и то, что по сравнению с A. astacus он быстрее растет, отличается большей плодовитостью и способностью питаться круглосуточно, а не только ночью, как широкопалый рак.
На территории Британских островов было замечено резкое сокращение ареала обыкновенной белки (Sciurus vulgaris) после завоза из Северной Америки близкого к ней вида Каролинской белки (Sciurus carolinensis), хотя природа конкурентного вытеснения осталась неизвестной. Виды, обитающие на островах, особенно страдают от вселенцев с материка, которые, как правило, оказываются более конкурентоспособными. Как отмечает Э. Майр (1968), большинство видов птиц, исчезнувших за последние 200 лет, были островными.
Очевидно, что увеличение области распространения одного вида, совпадающее с одновременным сокращением области распространения другого экологически близкого вида, вовсе не обязательно должно быть следствием конкуренции. На подобный сдвиг границ зон обитания могут влиять также иные биотические факторы, например деятельность хищников, доступность кормовых объектов или изменение абиотических условии. Так, в качестве примера конкурентного вытеснения рассматривалось ранее изменение распространения на Ньюфаундленде двух видов зайцев: полярного зайца (Lepus arcticus) н американского зайца-беляка (Lepus americanus). Более ста лет тому назад на острове обитал только полярный заяц, который населял самые различные биотопы, как в горах, так и в лесных долинах. Завезенный на остров в конце прошлого века заяц-беляк распространялся по лесным долинам, а полярный заяц стал встречаться только в горных безлесных районах. Была предложена простая гипотеза конкурентного вытеснения одного вида другим, но затем выяснилось (Bergerud, 1967), то в исчезновении полярного зайца из лесных районов виновен хищник — рысь (Lynx lynx), численность которой резко возросла после вселения на остров зайца-беляка. Косвенным доводом в пользу того, что пресс хищников сыграл в данном случае решающую роль, служит исчезновение полярного зайца из тех районов, куда не проник заяц-беляк, но которые по характеру растительности удобны для преследования зайцев рысью. Таким образом, гипотеза конкурентного исключения в данном случае, хотя и не была отвергнута полностью, должна была уступить гипотезе, учитывающей взаимоотношения трех видов: двух потенциальных конкурентов и одного хищника.