

Колебания системы хищник—жертва
Еще в 20-х гг. А. Лотка (Lotka, 1925), а несколько позднее независимо от него В. Вольтерра (1976) предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки—Вольтерры. Если предположить, что популяция жертв в отсутствие хищника растет экспоненциально, а пресс хищников тормозит этот рост, причем смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций), то мгновенная скорость изменения численности популяции жертв dN1/dt может быть выражена уравнением — dN1/dt = r1N1 – p1N1N2, где r1 — удельная мгновенная скорость популяционного роста жертвы, p1 — константа, связывающая смертность жертв с плотностью хищника, a N1 и N2 — плотности соответственно жертвы и хищника. Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости (которая в свою очередь зависит от интенсивности потребления хищником жертв) и постоянной смертности:
![]()
где р2. — константа, связывающая рождаемость в популяции хищника с плотностью жертв, а d2 — удельная смертность хищника (принимаемая постоянной). Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т. е. рост числа жертв лимитируется прессом хищников, а рост числа хищников — недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода из-под контроля хищника популяции жертв, хотя на самом деле такое бывает достаточно часто.
Н
Рис. 45. Простая модель, показывающая
возникновение устойчивых колебаний
численности хищника и жертвы: а —
положение изоклин, характеризующих
условие постоянства численности хищника
и жертвы. Стрелками показаны векторы
изменения в соотношении численности
хищника и жертвы; б — циклические
колебания численности хищника (пунктир)
и жертвы (сплошная линия), возникающие
согласно данной модели
Одна из первых попыток получения
колебаний численности хищника и жертвы
в лабораторных экспериментах принадлежала
Г. Ф. Гаузе (Gause, 1934). Объектами этих
экспериментов были инфузория парамеция
(Paramecium caudatum) и хищная инфузория
дидиниум (Didinium nasutum). Пищей для
парамеции служила регулярно вносимая
в среду взвесь бактерий, а дидиниум
питался только парамециями. Данная
система оказалась крайне неустойчивой:
пресс хищника по мере увеличения его
численности приводил к полному истреблению
жертв, после чего вымирала и популяция
самого хищника. Усложняя опыты, Гаузе
устраивал убежище для жертвы, внося в
пробирки с инфузориями немного стеклянной
ваты. Среди нитей ваты могли свободно
перемещаться парамеции, но не могли
дидиниумы. В таком варианте опыта
дидиниум съедал всех парамеций, плавающих
в свободной от ваты части пробирки, и
вымирал, а популяция парамеции затем
восстанавливалась за счет размножения
особей, уцелевших в убежище. Некоторого
подобия колебаний численности хищника
и жертвы Г. Ф. Гаузе удалось добиться
только в том случае, когда он время от
времени вносил в культуру и жертву и
хищника, и митируя
таким образом иммиграцию.
митируя
таким образом иммиграцию.
Ч
Рис. 46. Колебания численности двух
инфузорий — Paramecium aurelia (1)
и Didinium nasutum (2) —
в условиях разной обеспеченности кормом
Р. aurelia: а —
максимальное количество пищи; б —
минимальное количество пищи (по
Luckinbill, 1974)
М
Рис. 47. Изоклины стационарных
(сохраняющих постоянную численность)
популяций жертвы (а) и хищника (б) (по
Rosenzweig, MacArthur, 1963)
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 47, б, определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.
П
Рис. 48. Возникновение разных колебательных
режимов в системе хищник— жертва в
зависимости от взаиморасположения
изоклины стационарной численности
жертвы и хищника: 1
— область возрастания численности
жертвы; 2 — область возрастания
численности хищника (по Rosenzweig, MacArthur,
1963)
И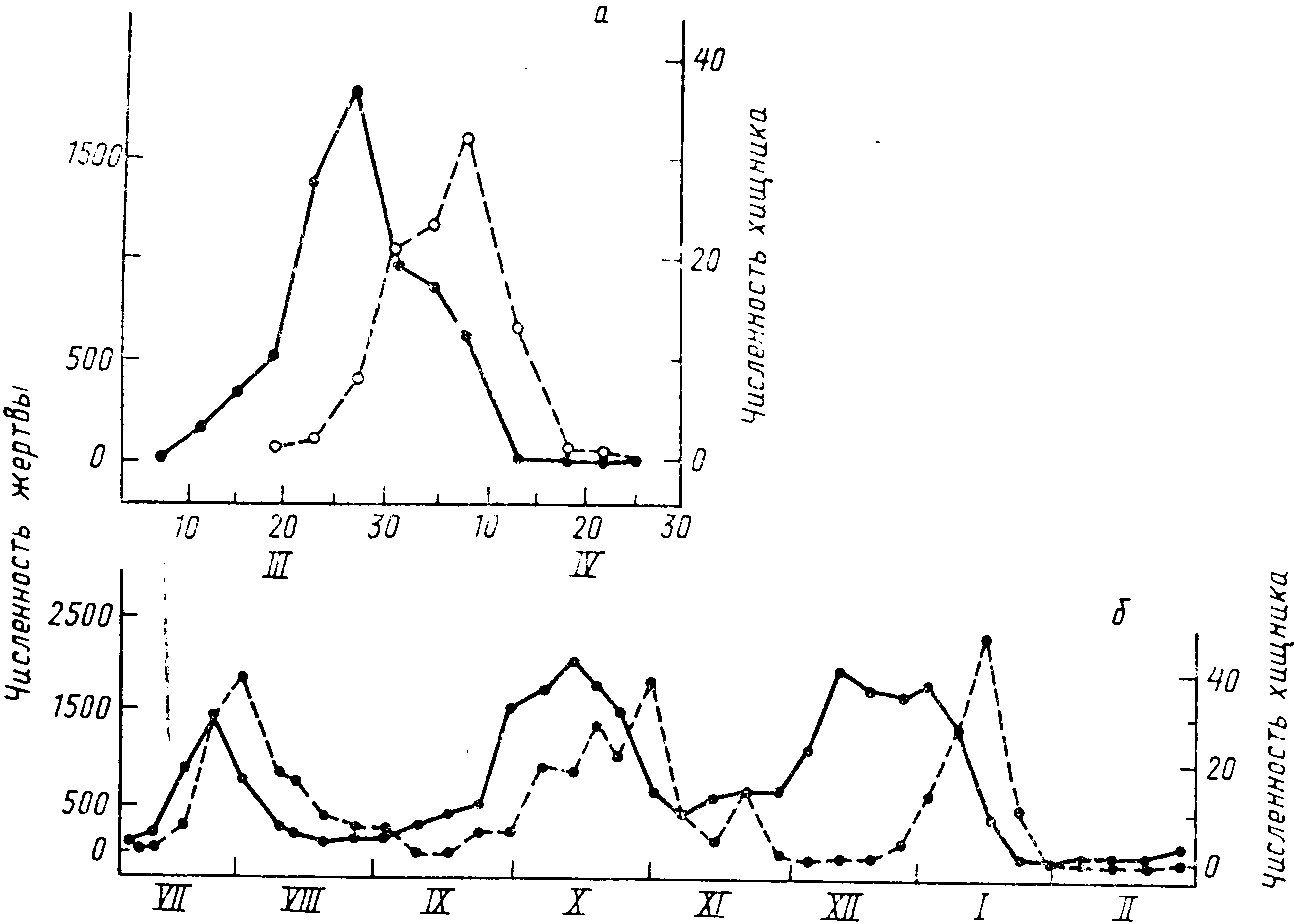 ными
словами, затухающие колебания соответствуют
ситуации, при которой хищник ощутимо
воздействует на популяцию жертв,
достигнувшую только очень высокой
плотности (близкой к предельной), а
колебания возрастающей амплитуды
возникают тогда, когда хищник способен
быстро увеличивать свою численность
даже при невысокой плотности жертв и
таким образом быстро ее уничтожить. В
других вариантах своей модели М.
Розенцвейг и Р. Мак-Артур показали, что
стабилизировать колебания хищник —
жертва можно, введя «убежище», т. е.
предположив, что в области низкой
плотности жертв существует область,
где численность жертвы растет независимо
от количества имеющихся хищников.
ными
словами, затухающие колебания соответствуют
ситуации, при которой хищник ощутимо
воздействует на популяцию жертв,
достигнувшую только очень высокой
плотности (близкой к предельной), а
колебания возрастающей амплитуды
возникают тогда, когда хищник способен
быстро увеличивать свою численность
даже при невысокой плотности жертв и
таким образом быстро ее уничтожить. В
других вариантах своей модели М.
Розенцвейг и Р. Мак-Артур показали, что
стабилизировать колебания хищник —
жертва можно, введя «убежище», т. е.
предположив, что в области низкой
плотности жертв существует область,
где численность жертвы растет независимо
от количества имеющихся хищников.
С
Рис. 49. Колебания численности
растительноядного клеща Eotetranychus
sexmaculatus и нападающего на него хищного
клеща Typhiodromus
occidentalis в лабораторной системе на лотках
с апельсинами и резиновыми мячиками: а —
система из 40 лунок (20 — с апельсинами);
б — система из 120 лунок (6 — с
апельсинами), усложняющая передвижение
хищника и облегчающая расселение
жертвы. Численность приведена как
среднее количество клещей, приходящихся
на один апельсин (по Huffaker, 1958)
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом б апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 мес., продемонстрировав три полных цикла колебаний численности (рис. 49, б). Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum), завезенная в Австралию, значительно снизила численность кактуса опунции46, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. Обычно эти бабочки откладывают яйца не на каждое попадающееся растение опунции, а на некоторые кусты, показавшиеся им в силу каких-то причин особо привлекательными. Вылупившиеся личинки при высокой плотности обычно полностью уничтожают свое кормовое растение, при этом много личинок гибнет, будучи не в состоянии расселиться на соседние кусты (если только они не расположены друг к другу ближе, чем на 2 м). В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию. Приведенный выше пример — еще одно свидетельство важности учета расселения и вообще динамики пространственного распределения популяции при изучении динамики ее численности. Фактически эти два процесса в ряде случаев должны рассматриваться вместе как две стороны одного процесса — движения численности в пространстве — времени.
Г
Рис. 50. Колебания численности зайца
(Lepus
americanus — 1) и рыси
(Lynx lynx — 2) в Канаде
по материалам заготовки шкур компанией
Гудзон-Бей (по MacArthur, Connel), 1966)