


22 :: 23 :: 24 :: 25 :: Содержание
11.4.3. Перемена направления движения ресничек
Большинство ресничек эпителия совершают биения в строго определенном направлении, однако реснички простейших и эпителия личинок оболочников способны менять направление рабочего гребка.
Наиболее широко исследованы механизмы поворота ресничек у простейших. Существует непосредственная связь между изменением направления гребка реснички и изменением направления движения метахронных волн. Другими словами, смещение направления рабочего гребка сопровождается аналогичным смещением направления метахронизма. Такого рода зависимость была использована для установления корреляции между направлением биений жгутиков и величиной мембранного потенциала у жгутикового простейшего Opalina. Для этого,
22
Рис. 11-22. Метод изучения нервной регуляции биения ресничек. При электрической стимуляции нерва, подходящего к жабрам мидии (Mytilus). реснички прекращают биения. А. Экспериментальная установка.
Изображение метахронных волн биения ресничек движется вдоль продольной оси щели, за которой с постоянной скоростью протягивается кинопленка. Б. Движения волн оставляют на пленке диагональный рисунок, который прерывается при изменении активности ресничек (В). В ответ на стимуляцию нерва на кинопленке появляется пробел, связанный с остановкой биений ресничек. В левом нижнем углу видна калибровочная отметка, соответствующая длине 20 мкм. (Takahashi. Murakami. 1968.)
варьируя концентрацию KCI во внешней среде, изменяли величину мембранного потенциала клетки. Затем наносили на график полученные величины мембранного потенциала и соответственно положение метахронных волн (рис. 11-23). По мере увеличения деполяризации мембраны направление рабочего гребка органеллы смещалось по часовой стрелке до тех пор. пока не достигало верхнего положения. У свободно плавающей клетки такая
переориентация биений органеллы приводит к изменению направления движения клетки на обратное. Описанное смещение ориентации рабочего гребка называют обращением хода реснички (или жгутика). У жгутиконосца Opalina, а также у Paramecium и большинства других инфузорий поворот ресничек лежит в основе реакции избегания, возникающей при появлении препятствия или раздражителя (см. разд. 8.7). Ориентация рабочего гребка меняется постепенно по мере увеличения уровня деполяризации мембраны, что несколько напоминает градуальную зависимость между силой мышечного сокращения и величиной мембранного потенциала.
Для эффективной работы механизма, вызывающего перемену направления рабочего гребка реснички, как и для мышечного сокращения, требуется повышенное содержание свободного внутриклеточного кальция. Реснички меняют направление своих биений лишь в том случае, если имеется внеклеточный кальций, и он проникает в клетку при деполяризации мембраны. Высокая чувствительность ресничного аппарата клетки к ионам кальция была продемонстрирована на экстрагированных с помощью детергента Paramecium (Naitoh, Kaneko, 1972). Мембраны клеток лишали свойств диффузионного барьера путем воздействий на них нейонным детергентом тритон Х-100, затем инфузорий вновь активировали, помещая в раствор, содержащий АТР и Са 2+ , где у них восстанавливалась способность плавать. Если концентрацию Са 2+ в реактивирующем растворе поддерживали на уровне ниже 10-6 М, клетки плыли вперед. Если же концентрация Са2+ превышала указанный уровень, реснички изменяли направление своего биения на противоположное, заставляя клетку двигаться назад (рис. 11-24). Эти результаты позволяют предположить, что направление рабочего гребка ресничек в интактной клетке также регулируется концентрацией внутриклеточного Са2+.
23
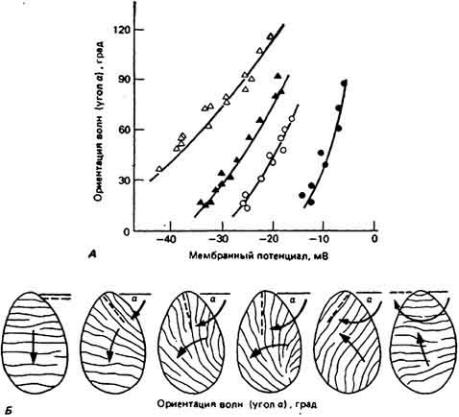
Рис. 11-23. Ориентация метахронных волн в зависимости от величины мембранного Потенциала. Для создания на мембране различных потенциалов жгутиковое простейшее Opatina погружали в растворы с разной концентрацией КСl. Потенциалы регистрировали с помощью микрозлектрода. и наблюдаемую при этом ориентацию метахронных волн (угол а) наносили на график соответственно величинам мембранных потенциалов (А). Чем больше была величина деполяризации мембраны, тем сильнее происходил разворот
метахронных волн в сторону переднего конца жгутиконосца. Результаты, полученные на отдельных организмах, обозначены на графике разными символами. Б. Направление движения волн по поверхности Opalina в зависимости от деполяризации, распространяющейся слева направо. (Kinosita. 1954.)
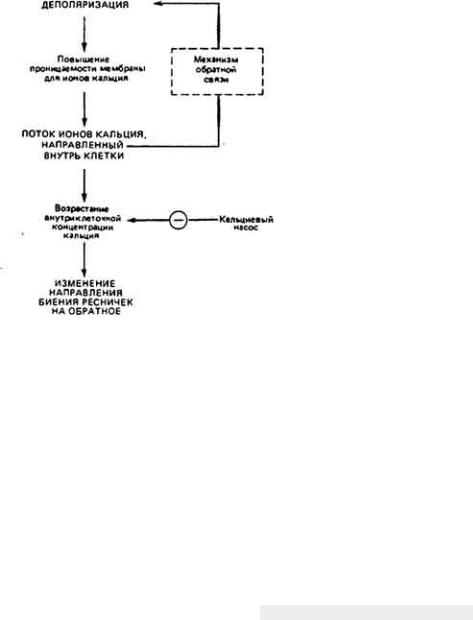
С помощью электрофизиологических экспериментов показали, что деполяризация клеточной мембраны у Paramecium приводит к активации кальциевых каналов. Результирующий входной ток Са 2+, который можно регистрировать с помощью электроизмерительных приборов, имеет характер импульса (рис. 11-25), но в отличие от нервного импульса он коррелирует по величине с интенсивностью приложенного стимула, а не следует закону "все или ничего". Входной ток ионов кальция через каналы в поверхностной мембране вызывает повышение его концентрации внутри ресничек. Ионы кальция, содержащиеся внутри реснички, влияют на скольжение микротрубочек, вызывая в конечном счете сдвиг в ориентации биений органеллы. Следовательно, сопряжение стимулируемой деполяризации мембраны с изменением направления биения ресничек обеспечивается у Paramecium входным током Са2+ (рис. 11-26).
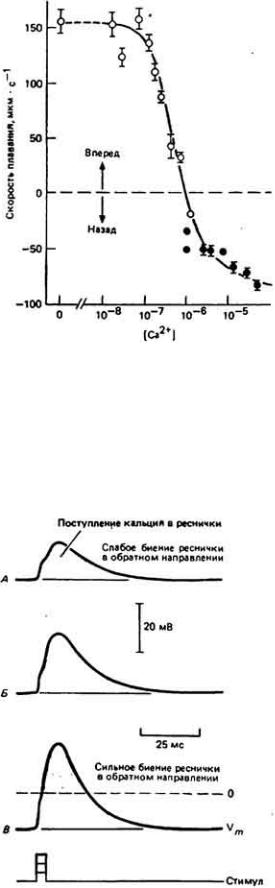
Рис. 11-24.
Зависимость между концентрацией Со2+ и направлением биений ресничек в экстрагированных с помощью детергента тритон X-100 препаратах Paramecium. При реактивации в растворе Мg-АТР экстрагированные клетки начинали плавать.
Если концентрация С2+ была ниже 10-6 М. клетки двигались вперед, если выше
10-6 М - назад. (Naiioh. Kaneko. 1972.)
24
Рис. 11-25.
Реакция инфузории Paramccium на добавление кальция. Короткие импульсы тока трех различных интенсивностей (линии внизу) вызывают увеличение реакции (А - В). Окрашенные области указывают примерную степень деполяризации,
возникающей за счет поступления в клетку ионов Са2+. На постепенное
увеличение входного тока Са2+ реснички реагируют изменением направления биений, сила и продолжительность которых коррелируют с величиной притока
Са2+. (Eckert. 1972.)
Рис. 11-26.
Сопряжение между поворотом реснички и величиной мембранного потенциала. Деполяризация при стимулировании мембраны заставляет открываться
кальциевые каналы. Благодаря большому электрохимическому градиенту Са 2+ поступает через эти каналы внутрь клетки, главным образом в реснички, поскольку большинство канальциевых каналов расположены в той части
мембраны, которая покрывает аксонему. Указанный входной ток Са 2+ внешне проявляется как слабо регенеративная деполяризация, вроде образования потенциала действия. Увеличение содержания свободных ионов кальция в
ресничках вызывает перемену направления рабочего гребка. Удаление Са2+ из реснички посредством транспорта через мембрану или диффузии в остальную часть клетки сопровождается возвращением к прежнему режиму биений. (Eckeri. 1972.)
25
22 :: 23 :: 24 :: 25 :: Содержание