

9 :: 10 :: 11 :: Содержание
11.1.5. Амебоидное движение
Рассматривая в обычный микроскоп крупных свободноживущих простейших
Amoeba proteus и Chaos carolincnsis, можно отметить у них единственную форму передвижения - амебоидную. Вакуолизированная сердцевина цитоплазмы, называемая эндоплазмой, устремляется вперед (в направлении движения простейшего) в одну или несколько псевдоподий (ложноножек), которые выпячиваются по мере постепенного, перетекания в них содержимого клетки (рис. 11-5). Ток эндоплазмы ограничен рамками периферической оболочки, выполненной из относительно гладкой, плотной и гелеподобной цитоплазмы (эктоплазмы). Она образуется на кончиках псевдоподий и действует как чехол, внутри которого
9
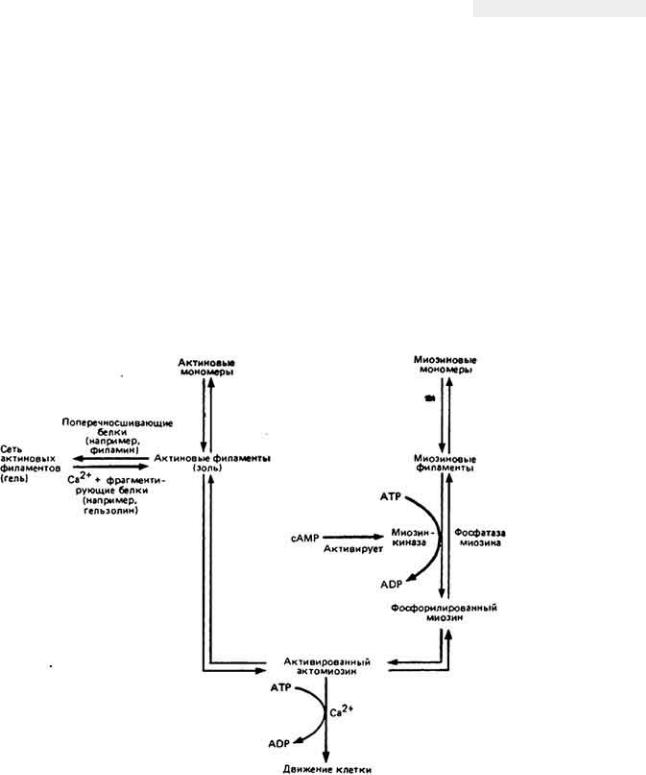
Рис. 11-4. Актин-
миозиновые взаимодействия, которые, по мнению исследователей, обеспечивают подвижность фибробластов и других амебоидных клеток. Участок, где происходит регуляция процесса, показан в цвете.
менее вязкая эндоплазма движется вперед. По ходу своего продвижения эндоплазма постепенно сгущается. Когда амеба перемещается с места на место, направление ее движения часто меняется. Чтобы изменить курс движения на противоположный, она перетекает в одну из своих псевдоподий, останавливается, затем втягивает се, одновременно перетекая в другую или образуя новую псевдоподию.
Один из извечных вопросов в отношении амебоидного движения - в каком месте продвигающейся клетки возникает движущаяся сила: выталкивается ли
эндоплазма вперед при сокращении позади себя эктоплазмы, подобно выдавливанию зубной пасты из тюбика, или же эндоплазму тянут вперед силы, возникающие в самой псевдоподии? Распределение сил внутри передвигающихся амебоидных клеток еще полностью не выяснено и, возможно, что на самом деле у разных типов клеток оно не одинаково. Проведенные с использованием оптических средств опыты на A. proteus и С. carolinensis (D. Lansing Taylor et al., 1978) привели к созданию модели молекулярных процессов, обусловливающих амебоидное движение у данных микроорганизмов. Сущность модели состоит в том, что актиновые фибриллы, поперечносшитые между собой с помощью желатинирующего фактора, например филамила, образуют очень вязкий гель (рис. 11-6, А, слева). Для того чтобы взаимодействовать с миозином и вызывать при скольжении филаментов сокращение структур клетки, гелсобразный актин должен превратиться в отдельные филаменты путем либо разрыва поперечных сшивок, либо фрагментации самих актиновых нитей. Подобное состояние вещества, лишенного сшивок между филаментами и имеющего низкую вязкость, называют золем. В этом состоянии отдельные актиновые филаменты могут реагировать с миозиновыми, обеспечивая сокращение при скольжении друг относительно друга (рис. 11-6, Б, справа). Сократительные силы передаются дальше в соседние области клетки через филаменты, соединенные с окружающим их гелеобразным актином. Для перехода актинового геля в золь необходимо повышенное содержание в цитоплазме свободного Са2+ или Н+. В присутствии миозина скольжение сократительных филаментов друг относительно друга возникает при повышении концентрации цитоплазматического Са 2+ и наличии источника энергии в виде АТР. Согласно
10
Рис. 11-5. Микрофотография гигантской амебы Chaos carolinensis, полученная с помощью светового микроскопа. Эндоплазма перетекает к псевдоподиям (показано стрелками). Обратите внимание на то, как сморщена втягиваемая задняя часть амебы (слева) по сравнению с гладкой поверхностью вновь образующихся псевдоподий. (С любезного разрешения К. Д. Аллен.)

приведенной схеме, сокращение в геле невозможно, поскольку актиновые нити в этом случае поперечно сшиты между собой и не в состоянии скользить вдоль миозина, развивая силу сокращения. Для того чтобы сокращение произошло, прежде всего необходим разрыв поперечных сшивок актиновых филаментов, который осуществляется при повышении концентрации Са2+ в цитоплазме.
11
9 :: 10 :: 11 :: Содержание

12 :: 13 :: 14 :: 15 :: Содержание
11.2.1. Типы движения
Существует много типов движения, обусловленных биениями, типа волновых изгибов или бегущих волновых изгибов. Легче всего описать те формы движения, при которых весь цикл волнообразного изгиба идет в одной плоскости пространства. Подобное движение, свойственное ресничкам некоторых типов клеток, было изучено с помощью высокоскоростной киносъемки (рис. 11-9). Цикл биения реснички можно условно разделить на две части -рабочий гребной (эффективный) удар вперед и восстановительное
движение. Во время рабочего гребка ресничка, оставаясь в целом относительно прямой, сгибается у своего основания по ходу движения в воде. При восстановительном движении реснички изгиб у основания начинает расширяться я передвигается вдоль органеллы по мере наклона ее базальной части в направлении, противоположном рабочему гребку. В результате ресничка, возвращаясь в исходное положение, необходимое для следующего гребка, затрачивает минимальное усилие на преодоление сопротивления внеклеточной окружающей среды. При таком движении реснички результирующий вектор силы, приложенной к внеклеточной среде, направлен параллельно поверхности клетки.
Двумерно-пространственное движение, т.е. движение в одной плоскости, характерно для ресничек эпителия многоклеточных организмов, особенно в тех случаях, когда длинные, тесно сгруппированные, частые и многочисленные реснички функционируют вместе, образуя циррус (обнаружен в жабрах двустворчатых моллюсков) или гребные пластинки (у гребневиков). Реснички простейших и некоторых типов эпителиальных клеток совершают более сложные биения не в одной плоскости, а в трехмерном пространстве. В этом случае во время рабочего гребка ресничка движется практически в одной плоскости, но во время восстановительного хода она выходит из нее, сильно наклонившись в одну сторону вблизи поверхности клетки (рис. 11-10).
Жгутикам свойственны разные формы движения: у одних оно имеет вид плоских волн, у других - спиральных вращений (рис. 11-7). С первого взгляда
12
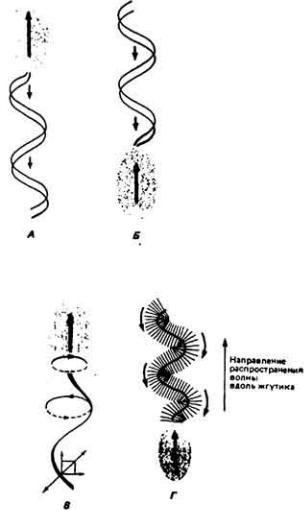
Рис. 11-7. Примеры движения жгутиков у различных
простейших. А, Б. Плоские водны. Клетка движется в направлении, противоположном распространению волны вдоль жгутика. У некоторых видов простейших жгутик расположен спереди (см. Б). В.
Спиральные волны создают более сложный комплекс векторов сил. добавляя при этом вращение тела клетки в направлении, противоположном спиральному вращению жгутика. Г. Перпендикулярно поверхности жгутика у Mastigamoeba setosa расположены похожие на волоски филаменты-мастигонемы, которые меняют направление развиваемой тяги на противоположное, продвигая клетку в ту же сторону, куда распространяется волна изгибов жгутика. Такое реверсирование тяги осуществляется благодаря тому, что мастигонемы, выступающие на внешней кривизне бегущих волнообразных изгибов органеллы, поворачиваются в направлении, обратном курсу следования волн жгутика (обозначено кривыми стрелками). (A: John el al. 1964; Б, В: Crell. 1973; Г: Jhan. Votta, 1972.)
может показаться, что волнообразные движения жгутика представляют собой пассивное распространение волн изгибов, возникающих под действием активных колебаний у основания органеллы, т. е. по
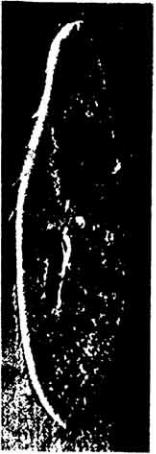
Рис. 11-8. Микрофотография плывущей Paramecium multimicronucleatum, полученная с помощью интерференционного микроскопа. Видны волны движения ресничек. Стрелками показаны реснички, производящие рабочий гребок назад (в нижней части рисунка), заставляя тем самым клетку двигаться вперед. Около × 700. (Machemer. 1972.)
аналогии с механизмом движения хлыста. На самом деле это не так: силы изгиба возникают по всей длине жгутика или в той его части, которая совершает движение. О том, что процесс идет именно так, говорят следующие факты.
1.Если бы активные движения у основания жгутика создавали пассивные волны изгиба, амплитуда этих волн постепенно снижалась бы по мере их продвижения к свободному концу органеллы, так как вязкость и плотность водной среды высоки в сопоставлении со скоростью движения жгутика (см. дополнение 16-3). Как видно на рис. 11-11, у бегущих волн, генерируемых в жгутиках спермиев морского ежа, такого снижения амплитуды нет.
2.Изолированные жгутики способны самостоятельно двигаться, несмотря на отсутствие естественного контакта с клеткой.
13

Рис. 11-9. Анализ движения гигантской составной реснички, обнаруженной в жабрах мидии, с помощью высокоскоростной (450) кадров в секунду) киносъемки. Нумерация кадров дана внизу. На рис. А и Г показан каждый восьмой кадр, на рис. Б - каждый четвертый, на рис. В-каждый второй. Рабочий гребок начинается на 40-м кадре и заканчивается примерно на 84-м. Востановительное движение продолжается приблизительно от 84-го по 148-й кадр. Прежде чем совершить после восстановительного движения новый гребок, ресничка на некоторое время распрямляется. Движения данной реснички происходят главным образом в одной плоскости, и поэтому они проще, чем трехмерные движения, присущие ресничкам простейших. (Ваbа. Hiramoto, 1970.)
3.У некоторых жгутиков волны изгиба возникают на свободном конце органеллы и распространяются по направлению к ее основанию (рис. 11-7, Б).
4.Движение некоторых жгутиков ограничено небольшим участком на свободном конце. Проксимальная же область жгутика остается неподвижной и распрямленной.
Рис. 11-10. Воспроизведение стадий цикла биения реснички у инфузории Paramecium. Показано семь положений, пронумерованных в порядке следования движений, характерных для отдельной реснички.
Рабочий гребок (стадии 1-3) идет в одной плоскости. Восстановительное движение (стадии 4-7)

осуществляется вблизи поверхности клетки, причем сама ресничка наклонена все время в одну сторону. Короткой стрелкой обозначено направление хода волны. Положение передней части тела простейшего указано cmpелкой с надписью "Передн.". Модель построена по микрофотографиям биений ресничек.
(Machemer, 1972).
Рис. 11-11. Форма биений жгутика при движении сперматозоида морского ежа. А. На многократных стробоскопических снимках видно, что у бегущих волн изгибов жгутика амплитуда остается постоянной на всем протяжении органеллы. Б. Распрямленные участки аксонемы чередуются с куполообразными. (Brokaw, 1965.)
14
Рис. 11-12. Иерархические уровни внутреннего строения аксонемы. Тубулиновые гетеродимеры собираются в протофиламенты - строительные блоки, из которых состоят микротрубочки. Основная часть хвоста спермия представляет собой аксонему. окруженную поверхностной мембраной. (По N. Wessels. How Living Cells Shape. Copyright © 1971. Scientific American. Inc. AH rights reserved.)
15
12 :: 13 :: 14 :: 15 :: Содержание