


44 :: 45 :: Содержание
12.3.5. Канальцевая секреция
Нефрон имеет несколько четко различимых специализированных систем, которые секретируют вещества в просвет канальца посредством их переноса из плазмы крови. Наиболее изучены те системы, которые отвечают за секрецию К+, Н+, NH3, органических кислот и органических оснований. Казалось бы, число секреторных механизмов и транспортных молекул должно быть ограничено; однако
44
на самом деле нефрон способен сегрегировать бесчисленное количество "новых" веществ, включая лекарственные препараты и токсины, а также эндогенные, обычно присутствующие в организме молекулы. Каким же образом нефрон узнает и переносит из крови все эти столь разнообразные вещества? Повидимому, у позвоночных это связано с функцией печени, обеспечивающей модификацию многих из указанных молекул, таким образом, чтобы они могли реагировать с транспортными системами, расположенными в стенке нефронов. Подобные секреторные механизмы важны для организма потому, что они удаляют из крови потенциально опасные вещества. В печени многие из этих веществ наряду с обычными метаболитами конъюгируют с глюкуроновой кислотой или сульфатом. Полученные таким путем два типа конъюгированных молекул активно переносятся системой, которая узнает и секрeтирует органические кислоты. Поскольку соответствующие конъюгированные молекулы имеют высокую полярность, то, перенесеные с помощью транспортного механизма в просвет нефрона, они уже не могут легко диффундировать через стенку нефрона обратно в околоканальцевое пространство, а оттуда поступить в кровь. В итоге данные вещества экскретируются с мочой.
12.3.5.1. Секреция йонов калия
Большая часть легко фильтрующихся в клубочках ионов калия обычно реабсорбируется из фильтрата в проксимальных канальцах и петлях Генлe. Скорость активной реабсорбции в канальце и петле не снижается даже в том случае, когда концентрация К + в крови и фильтрате сильно возрастает в ответ на избыточное потребление организмом этого иона. Однако дистальные канальцы и собирательные протоки способны не только реабсорбировать, но и секретировать ионы калия. Секретируя калий, данные структуры стремятся достичь ионного гомеостаза в случае поступления в организм необычайно большого количества этого металла. Транспорт К+, по-видимому, зависит от его постуления в клетки канальцев из тканевой жидкости, обусловленного активностью обычного Nar+ - Ka+-насоса (рис. 12-16), с утечкой К+ из цитоплазмы в канальцевую жидкость. Калий может просто диффундировать по электрохимическому градиенту из клеток почечных канальцев в просвет,
потому что канальцевая жидкость электроотрицательна по отношению к цитоплазме. Секреция К+ с помощью данных механизмов стимулируется адренокортикальным гормоном-альдостероном, который высвобождается в ответ на повышение содержания К+ в плазме крови.
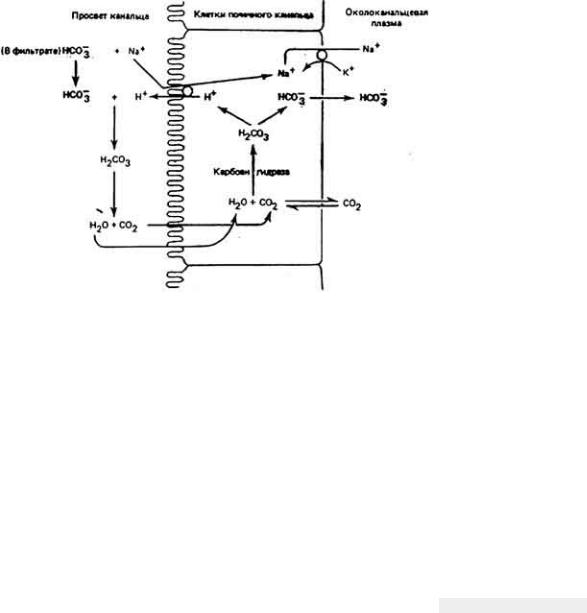
Рис. 12-16.
Реабсорбция из фильтрата ионов бикарбоната и натрия. Внутриклеточный фермент карбоангидраза катализирует образование H2CO3 из СO2 и воды.
Свободные ионы водорода поступают в просвет канальцев в результате
противоточного транспорта Na+ -H+. В просвете канальца H+ взаимодействуют с отфильтрованными в клубочках из плазмы ионами HCO3 и
образуют H2СO3 - соединение, которое затем диссоциирует на Н2O и СO2. Оба
этих вещества диффундируют обратно в клетки почечных канальцев чтобы вновь повторить цикл химических превращений. Натрий поступает из просвета канальца в клетку по концентрационному градиенту, обмениваясь при этом "один
к одному" с ионами H+. Затем натрий выкачивается из клетки в области серозной поверхности под действием Na|+ -К+-насоса. Общий смысл данных
превращений -вернуть в организм бикарбонат-ион и Na+, так как в противном случае эти ионы были бы выведены с мочой. (Pitts, 1974; Vonder. 1980.)
45
44 :: 45 :: Содержание

46 :: 47 :: 48 :: Содержание
12.3.6. Регуляция почками кислотно-основного состояния
Мембрана, выстилающая просвет почечных канальцев, имеет противоточную транспортную систему, с помощью которой ионы Н+ удаляются из цитоплазмы эпителиальных клеток в просвет канальца в обмен на ионы N+. Данный процесс происходит в проксимальном канальце и далее на всем протяжении нефрона. В результате первичная моча, проходя по канальцу, становится все более кислой. Почечные клетки секретируют в просвет нефрона ионы Н+, тогда как ионы ОН- остаются в цитоплазме. Поступившие в просвет канальца ионы Н+ реагируют с
HCO - , образуя Н2СО3-соединение, которое диссоциирует на Н2О и СО2 (рис.
3
12-16). Несмотря на то что ионы Н+ заряжены положительно, в просвете канальца избыточного положительного заряда не возникает, так как протоны оттуда удаляются за счет противоточного транспорта, сопряженного с пассивным переходом Na+ по концентрационному градиенту из просвета нефрона в клетки почечного канальца. Таким способом движение Na+ в цитоплазму поддерживает противоположно направленное движение Н+ в просвет канальца. Тс ионы Na+, которые поступают этим путем в почечные клетки, затем удаляются из них с помощью АТР-зависи-мого Na + - К4 - насоса, локализованного в поверхностной мембране, обращенной в околоканальцевоe пространство. Как мы далее увидим, секреция Н+ в первичную мочу имеет особое значение для регуляции концентрации протонов в жидких средах организма.
Внеклеточную величину рН регулируют несколько буферных систем жидких сред организма. Все они находятся в состоянии равновесия друг с другом, насколько позволяет компартментация в тканях. Поэтому изменение буферной активности одной из данных систем вызовет сдвиг в концентрации Н + жидких сред, причем величина этого сдвига будет зависеть от всех остальных буферных систем. Однако, вне всякого сомнения, наиболее важной буферной системой, которая прежде всего отвечает за регуляцию рН во внеклеточном пространстве, у позвоночных является бикарбонатная буферная система:
Н2О + СО2 Н2СО3 Н+ + НСО - .
3
Эта система связана в первую очередь с дыхательным обменом, определяющим в основном концентрацию СО2 в организме. Например, когда дыхательный обмен уменьшается, а уровень СО2 возрастает, происходит снижение рН крови из-за накопления протоков в бикарбонат-ионов (см. разд. 14.6). Кроме того, данная буферная система связана с функцией почек, которые в конечном счете отвечают за поддержание концентрации НСО 3 в плазме на постоянном уровне.
На уровне отдельного нефрона концентрация бикарбоната в плазме крови регулируется двумя способами. Во-первых, за счет изменения скорости реабсорбции Н2СО3 из клубочкового фильтрата; во-вторых, за счет превращения СО2 и Н2О в бикарбонат, который затем направляется в плазму. Оба указанных
механизма, как мы увидим ниже, требуют активного переноса Н+ из клеток почечного канальца в его просвет.
12.3.6.1. Реабсорбция бикарбоната
Поскольку бикарбонат-ион (НСО3 ), как и все мелкие молекулы, свободно проходит из клубочка в канальцевую жидкость, он должен быть реабсорбирован для предотвращения развития тяжелого ацидоза из-за появления избытка кислоты в жидких средах организма вне нефрона. Действительно, в норме бикарбонат (вещество, обладающее щелочными свойствами) почти целиком реабсорбируется и в конечной моче почти отсутствует. Однако реабсорбция идет не по пути активного переноса НСО3 из просвета канальца. Вместо этого,
как мы уже отмечали ионы Н+, секретируемые в просвет, соединяются с бикарбонатом, образуя Н2СО3 (рис. 12 - 16). Кроме того, молекулы О2, находящиеся в клетках почечных канальцев, реагируют с водой в присутствии фермента карбоангидразы и тоже образуют Н2СО3. Последнее вещество сразу
же диссоциирует на Н+ и HCO - , причем анион диффундирует в
3
околоканальцевое пространство. Тем временем ионы Н+ соединяются с ионами Н-, которые освобождаются на поверхности мембраны в результате гидролиза НОН, происходящего в тот момент, когда Н + активно переносится через мембрану внутрь просвета канальца. Ионы водорода, поступившие в просвет канальца, замещают ионы Na+, пассивно диффундирующие из просвета в клетки канальца, и, соединившись с НСО3 , образуют СО2 и Н2О. Двуокись углерода, находящаяся в просвете канальца, которая без труда может проходить через клеточную мембрану, диффундирует в клетку, а затем поступает в плазму крови в виде СО2 или НСО3. Общий результат этих превращений состоит в том, что
НСО - и Na+ переходят из мочи в плазму крови, где они заменяют те
3
бикарбонат-ионы, которые покинули плазму во время клубочковой фильтрации. Обратите внимание на тот факт, что ионы водорода, активно секретируемые в просвет канальцев, не остаются в моче, а включаются в состав воды, большая часть которой реабсорбируется в нефроне до образования конечной мочи.
46
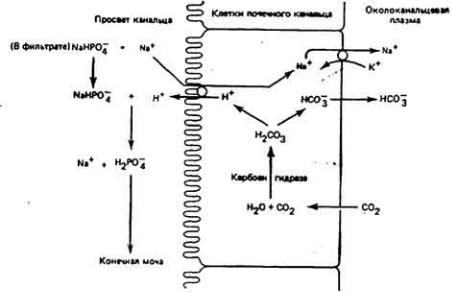
Рис. 12 - 17.
Экскреция ионов водорода. Протоны, продуцируемые и секретируемые, как показано на рис. 12 - 16. обмениваются с ионами натрия, которые переходят из фильтрата в клетки почечного канальца. Поступившие в просвет канальца ионы
H+ взаимодействуют с ионами фосфата, содержащимися в фильтрате, и
образуют ионы Н2РO - . которые не способны проникнуть сквозь клеточную
2
мембрану и поэтому остаются в моче. (Pins. 1974; Vender, 1980.)
12.3.6.2. Экскреция кислоты
Удаление из плазмы крови избытка водородных ионов - важная функция почек. Происходит это путем секреции Н+ в обмен на Na+, который, наоборот, поступает в клетки канальцев. Водородные ионы "удерживаются" в моче, как
правило, в виде заряженных молекул - NН + |
или Н РО - |
, которые не могут |
|
4 |
2 |
4 |
|
|
|
||
проходить сквозь клеточную мембрану. Одновременно в плазму крови
поступают новые ионы НСО - . Подобная система показана на примере
4
экскреции ионов Н+, связанных с фосфатом (рис. 12 - 17). Здесь Н2СО3
образуется тоже из СО2 и Н2О и диссоциирует на ион HCO - , поступающий в
3
плазму, и Н+ , который в это время активно переносится в просвет канальца.
Часть секретируемых ионов водорода соединяется с HPO |
2- ионом, который в |
|
|
4 |
|
клубочке свободно фильтруется из плазмы и образует |
Н 2 |
- . Последний не |
|
|
4 |
проникает сквозь мембраны канальцев, движется по нефрону без реабсорбции и в конечном счете выводится из организма с мочой. Следует отмстить, что в описанных процессах участвуют те же самые механизмы, что и в процессах реабсорбции бикарбоната, показанных на рис. 12-16. Различие, состоит в том, с каким анионом реагирует Н+ , будучи секретируемым в просвет канальца.
В просвете канальцев секретированный ион водорода реагирует также с
NH , образуя NH + |
(рис. 12 - 18). Как и Н РО - |
, |
NH - - ион с высокой |
||
3 |
4 |
2 |
4 |
|
4 |
|
|
|
|||
полярностью, который не в состоянии проникать сквозь клеточную мембрану. Он выходит из нефрона вместе с мочой без реабсорбции, унося с собой
секретированный протон. Аммиак - предшественик NH + -образуется в клетках
4
почечных канальцев при ферментативном дезаминировании аминокислот в отличие от фосфата, который попадает в канальцевую жидкость вследствие клубочковой фильтрации. Молекула аммиака свободно диффундирует сквозь клеточную мембрану в просвет канальца благодаря своему нeполярному и неионизированному состоянию. В конечном счете аммиак задерживается здесь, реагируя в просвете канальца с активно секретируемым Н+ и в результате превращаясь в ион аммония, для которого стенки канальцев непроницаемы. Таким образом, NH+ в моче служит транспортным средством для выведения из организма как атомов азота, так и водородных ионов. Секреция аммиака - процесс высоко адаптивный. У млекопитающих, впавших в состояние метаболического ацидоза (избыточная продукция и кислоты), наблюдают резкое увеличение синтеза и экскреции аммиака, потому что это основной адаптивный механизм в организме, корректирующий избыток кислот.
47
Рис. 12-18.
Секреция аммиака. NH3. который образуется в почечных клетках при
дезаминировании глутамина и аминокислот, свободно диффундирует из клетки в плазму и просвет почечных канальцев. В просвете аммиак реагирует с ионами водорода, секретируемыми почечными клетками; при этом образуется ион
аммония (NH - ), который не способен проникать через клеточные мембраны, т. е.
4
диффундировать обратно в клетку, и поэтому остается в моче. Подобный захват аммиака приводит к созданию крутого градиента концентрации аммония и облегчает тем самым его диффузию в просвет казкильцев. (Vander. 1980.)
48
46 :: 47 :: 48 :: Содержание

48 :: 49 :: 50 :: 51 :: 52 :: Содержание
12.3.7. Механизмы концентрировання мочи в нефроне
У птиц и млекопитающих моча становится концентрированной по причине осмотического выхода из нее воды-процесса, который происходит в собирательных протоках по ходу их в мозговом слое почки. Осмотический градиент, необходимый в собирательных протоках для удаления из мочи воды, устанавливается благодаря удивительному сочетанию клеточной специализации и анатомической организации структур в мозговом слое почки. У позвоночных существует четкая корреляция между строением почки и ее способностью продуцировать гипертоническую относительно жидких сред организма мочу. У всех животных, способных продуцировать гипертоническую мочу (т.е. у млекопитающих и птиц), почки имеют нефроны с характерной особенностью - петлей Генле. Последние научные данные позволяют предполагать, что почки пласти-но-жаберных рыб имеют подобное строение. Как правило, чем длиннее петля и чем глубже она заходит в мозговое вещество почки, тем сильнее концентрирующая способность нефрона. Поэтому почки млекопитающих, живущих в пустыне, имеют самую длинную петлю Генле и продуцируют наиболее гипертоническую мочу. Этот факт наряду с данными о постеленном нарастании тоничности внутри- и внеканальцевой жидкости по направлению к более глубоким зонам мозгового вещества почки (рис. 12-19) привел Аржитэ (В. Hargitay) и Куна (Werner Kuhn) в 1951 г. к предположению о том, что петля Генле действует как противоточная множительная система (дополнение 12-2).
Вначале проверка столь привлекательной и правдоподобной гипотезы представлялась весьма сложным делом из-за трудностей, связанных с отбором проб внутриканальцевой жидкости в тонких участках петли Генле. Измерение точки плавления жидкости в срезах замороженной почки позволило выявить рост осмотической активности как внутри, так и вне канальцевой жидкости в направлении от верхних к нижним концам петли Генле. Существующий
кортикомедуллярный градиент концентрации изображен в виде клина на рис. 12-20. В дальнейшем гипотеза о противоточном механизме концентрирования мочи проверялась методом искусственной перфузии in situ1 отдельных участков петли. Жидкость, поступающая из проксимального канальца в нисходящее колено петли, изоосмотична по отношению к внеклеточной жидкости в данном месте (в наружном слое мозгового вещества) и имеет концентрацию около 300 мосмоль-л-1. По мере движения жидкости вдоль нисходящего колена ее концентрация плавно возрастает и в области поворота петли
48
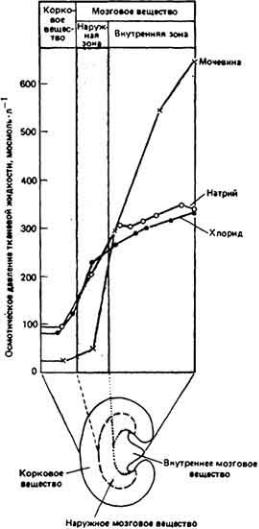
Рис. 12-19.
Распределение концентраций растворенных веществ вдоль оси почки млекопитающих. По мере продвижения от коры почки к глубокий зонам мозгового
вещества концентрация мочевины, Na+ и Сl- возрастает. Обратите внимание, что наибольшее увеличение концентрации мочевины происходит во внутреннем мозговом слое почки, тогда как наибольший подъем концентрации NaCl - в
наружном мозговом слое. Поскольку осмотические вклады Na + и Сl- суммируются, вклады молекул NaCl и мочевины в осмолярность глубоких зон мозгового вещества почки примерно равны между собой. (Ullrich et al.. 1961.)
Генле достигает (у большинства млекопитающих) 1000-3000 мосмоль- -1. В этом месте жидкость также почти изоосмотична относительно внеклеточной жидкости глубоко расположенных областей мозгового вещества почки. Далее мы кратко рассмотрим, почему тканевая жидкость глубоких зон мозгового вещества имеет столь высокую осмотическую концентрацию.
Канальцевая жидкость, опускаясь по нисходящему колену, вступает в осмотическое равновесие с тканевой жидкостью. Это происходит потому, что стенки нисходящего колена петли Генле относительно проницаемы для воды и гораздо менее проницаемы для NaCl и мочевины. Следовательно, концентрация калальцевой жидкости, вначале равная примерно 300 мосмоль • л ~', достигает 1000 -3000 мосмоль-л"1 (у некоторых пустынных млекопитающих даже больше), т.е. становится равной осмотической концентрации тканевой жидкости в том мете, где петля поворачивает (рис. 12-15). По мере движения жидкости
вверх вдоль толстого участка восходящего колена в ней из-за активного транспорта солей через стенку канальца усиливается потеря NaCl (но не воды). Как тонкий, так и толстый участки восходящего колена петли относительно непроницаемы для воды.
Функциональная асимметрия между нисходящим и восходящим коленом петли Генле в совокупности с принципом противотока объясняет наличие кортикомедудлярных1 градиентов концентрации NaCl и мочевины, показанных на рис. 12-19. Осмотическая потеря воды из нисходящего колена позволяет достичь в просвете этого колена равновесия с осмотической концентрацией тканевой жидкости. Поскольку первичная моча все время движется вдоль просвета канальца, она постоянно отдает воду в тканевую жидкость, осмотическая кондетрация которой растет по мере приближения к изгибу петли Генле (рис. 12-20).
Полагают, что интерстициальный осмотический градиент возникает при сочетании активного переноса NaCl из восходящего толстого участка петли и избирательной пассивной проницаемости стенки для воды, соли и мочевины в определенных участках нефрона. Вспомним, что проницаемость стенок нисходящего колена петли Генле высока для воды и низка для соли и мочевины, тогда как в восходящем колене, напротив, - высока для соли и низка для воды и мочевины. Начав рассмотрение явлений в нефроне со стадии 1 (рис. 12-21), мы видим, как Na+ и Сl-1 активно удаляются из просвета толстого участка восходящего колена петли Генле и дистального канальца. В результате дополнительного поступления NaCl в тканевую жидкость коры и наружного слоя мозгового вещества почки происходит осмотическая потеря воды (стадия 2). Вода выходит из тех же самых участков нефрона и из непроницаемого для соли нисходящего колена, расположенного в коре и наружной зоне мозгового вещества. Из-за общей потери воды и соли из петли Генле и последующей потери обоих веществ из дистального канальца в собирательном протоке
49
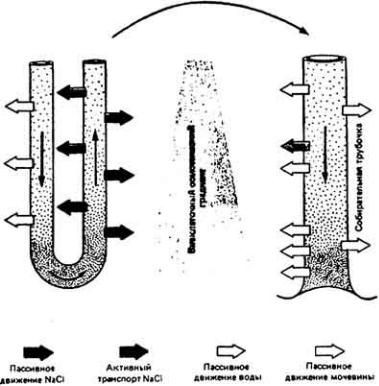
Рис. 12-20. Противоточная модель петли Генле. Места активного транспорта NaCl через клеточную мембрану нефрона показаны сплошными цветными стрелками. Градиент концентрации соли во внеклеточном пространтве изображен в виде клина серого цвета. Осмотическое движение воды в тканевую жидкость отмечено черными контурными стрелками; пассивная диффузия мочевины - цветными контурными срелками.
создается высокая концентрация мочевины. Важная особенность этого процесса состоит в том, что собирательный проток становится высокопроницаемым исключительно для мочевины в тех его отделах, которые глубоко заходят в мозговое вещество почки. Там мочевина выходит из протока по концентрационному градиенту (стадия 3), повышая этим осмотическую концентрацию тканевой жидкости во внутреннем слое мозгового вещества. Изза высокой осмотической концентрации тканевой жидкости возникает ток воды из нисходящего колена (стадия 4) с развитием у основания петли Генле очень высокой внутриканальцсвой концентрации растворенных веществ. Затем высококонцентрированная канальцевая жидкость движения вверх по тонкому участку восходящего колена, который имеет очень высокую проницаемость в отношении соли. Здесь из канальцевой жидкости начинает выходить NaCl (стадия 5), что снижает концентрационный градиент. Дальнейшие потери NaCl в толстом участке идут уже за счет процесса активного транспорта через мембрану клетки (стадия 1). Нижний отдел собирательного протокаединственный участок нефрона, обладающий высокой проницаемостью для мочевины. При этом небольшое количество мочевины, вновь поступившее в более проксимальные части нефрона, просто повторно вовлекается в цикл процесса. Высокая осмотическая концентрация тканевой жидкости внутреннего слоя мозгового вещества (рис. 12-19) обусловлена в основном пассивным накоплением здесь мочевины в результате работы противоточного концентрирующего механизма нефрона. Если бы восходящее колено петли было таким же проницаемым для мочевины, как и собирательный проток, то подобное накопление мочевины отсутствовало бы. Оно не имело бы места и в том случае, если бы не происходило активного удаления NaCl (вместе с

пассивно движущейся водой) и мочевина не начинала концентрироваться в собирательном протоке.
Интересно заметить, что градиент концентрации мочевины в мозговом веществе почки устанавливается благодаря работе главным образом пассивных механизмов, несмотря на особое значение такого функционального компонента противоточной системы, как активный перенос NaCl. На этот компонент расходуется большая часть энергии, необходимой для возникновения концентрационного градиента NaCl и мочевины. Результатом подобного сочетания специализации клеток и морфологии ткани
50
Рис. 12 - 21. Основные характеристики модели противоточного умножителя в почке. Кортикомедуллярный осмотический градиент возникает в результате активного накопления NaCl и пассивного накопления мочевины в мозговом веществе почки. Устойчивый градиент (рис. 12 - 19) зависит от неодинаковой проницаемости разных участков нефрона и активного транспорта соли через мембрану этих участков, а также от анатомического строения нефрона и его кровоснабжения (капилляры vasa recta не показаны). Размеры стрелок отражают относительную величину потоков веществ. Серым клином изображен осмотический градиент во внеклеточной жидкости. (По данным Jamison. Maffly. 1976.)
служит стационарный Кортикомедуллярный градиент концентрации мочевины и NaCl, при котором осмотическая концентрация как в канальцевой жидкости, так и в околоканальцевом пространстве непрерывно растет в направлении мозгового вещества почки. Этот градиент определяет конечный переход воды из собирательных протоков в тканевую жидкость за счет осмоса и последующее образование гиперосмотической мочи.
Между длиной петли Генле и способностью почек млекопитающих
концентрировать мочу существует четкая корреляция. Наибольшей длины петли Генле достигают у обитателей пустыни, например кенгуровой крысы. В длинных петлях на участке от коры до мозгового вещества возникают более высокие общие осмотические градиенты, обеспечивающие более эффективное всасывание воды из просвета собирательных протоков.
Характерной особенностью в противоточной организации системы кровоснабжения нефрона являются прямые сосуды (vasa recta). Их основная роль состоит в поддержании стационарного градиента концентрации веществ в тканевой жидкости. Кровь из коркового слоя почки опускается в глубокие зоны мозгового вещества по капиллярам, которые образуют вокруг каждого нефрона петлеобразные сети (рис. 12-7). Затем она поднимается в корковый слой. При такой системе циркуляции кровь с помощью осмотического механизма забирает соль и отдает воду. В итоге по мере поступления крови в глубь мозгового вещества тканевая жидкость, окружающая сосуды, становится все более насыщенной осмотически активными веществами. Противоположный процесс возникает, когда кровь возвращается в корковый слой и вступает в контакт с тканевой
51
жидкостью, осмотическая концентрация которой постепенно падает. Следовательно, осмотическая концентрация крови во время циркуляции по прямым сосудам изменяется ненамного, хотя вода и растворенные в ней вещества, рeабсорбированные из клубочкового фильтрата по мере его прохождения по нефрону, уносятся с кровью. Однако количество крови, циркулирующей по этим сосудам, составляет лишь небольшой процент от общего объема крови, омывающей почку.
Противоточное расположение прямых сосудов обеспечивает высокую скорость почечного кровотока (что особенно необходимо для эффективной клубочковой фильтрации) без нарушения устойчивости кортикомедуллярного градиента концентрации соли и мочевины. Скорость кровообращения в почках - действительно, одна из наиболее высоких скоростей кровотока, обнаруженных в тканях позвоночных. Когда кровь покидает клубочек и по прямым сосудам поступает в мозговое вещество, в нее пассивно переходят NaCl и мочевина из тканевой жидкости при непрерывно возрастающей осмотической концентрации последней. Максимальная концентрация NaCl и мочевины в крови достигается, когда кровь делает поворот в петле прямого сосуда, расположенной в глубине мозгового вещества. Поднимаясь обратно в корковый слой, кровь теряет избыток NaCl и мочевины, которые путем диффузии поступают обратно в тканевую жидкость, и покидает почку. Однако в действительности, прежде чем выйти за пределы почки, кровь забирает с собой некоторое количество воды, вышедшей из кровеносного русла во время клубочковой фильтрации. Это происходит из-за повышенного во время ультрафильтрации коллоидно-осмотического давления плазмы крови.
Итак, образование мочи начинается в проксимальном канальце с

концентрирования клубочкового фильтрата и превращения его в гиперосмотическую жидкость. В проксимальном канальце из первичной мочи реабсорбирует в осмотически эквивалентных количествах 75% соли и воды; мочевина и некоторые другие вещества остаются. Осмотическая концентрация первичной мочи, движущейся по петле Генле и дистальному канальцу, несколько изменяется. Однако противоточный механизм обеспечивает по ходу петли Генле устойчивый концентрационный градиент в тканевой жидкости мозгового вещества. Данный градиент создает основу для удаления воды из мочи осмотическим путем по мере продвижения жидкости вниз по собирательной трубочке, расположенной в мозговом веществе. Этот процесс идет без активного переноса воды через стенку нефрона.
52
1На месте нахождения.- Прим. перев.
1То есть между корковым и мозговым веществом почек.- Прим. перев
48 :: 49 :: 50 :: 51 :: 52 :: Содержание