


32 :: 33 :: 34 :: Содержание
12.1.1. Факторы, влияющие на облигатный осмотический обмен
1.Концентрационные градиенты между внеклеточными пространствами (компартментами) и окружающей средой. Чем больше разница между концентрацией вещества во внешней среде и в жидких средах организма, тем более выраженной становится направленность результирующей диффузии в сторону меньшей концентрации этого вещества. И если организм лягушки, погруженной в пресноводный водоем, стремится к обогащению водой за счет гипотонической окружающей среды, то морской костной рыбе, напротив, угрожает потеря воды из организма в гипертоническую окружающую среду.
2.Отношение поверхность - объем. Объем животного пропорционален кубу, а величина поверхности тела - квадрату его линейных размеров. Иными словами, у мелких животных отношение величины поверхности тела к объему будет больше, чем у крупных. Из сказанного следует, что у мелкого животного величина поверхности кожи, через которую вода и растворенные вещества могут обмениваться с окружающей средой, больше относительно количества содержащейся в теле жидкости, чем у крупного животного. Так, при одинаковой
результирующей скорости обмена через кожу (в моль-с -1 см-2) мелкое животное будет обезвоживаться (рис. 12 - 4) или, напротив, насыщаться водой быстрее, чем крупное животное с той же формой тела.
3. Проницаемость внешних покровов тела. Внешние покровы тела действуют как барьер между
32
Рис. 12-4. При одинаковой испарительной нагрузке, когда испарение воды обеспечивает поддержание бо.гее низкой температуры тела по сравнению с температурой окружающей среды, мелкие животные обезвоживаются быстрее, чем крупные, из-за присущей им высокой величины отношения площади поверхности те,\а к его массе. На данном графике показано количество воды, которое утрачивается организмом за час в жарких условиях пустыни (в процентах от массы тела). (Edney, Nagy. 1976.)
внеклеточными пространствами (компартментами) и внешней средой. Величина
проницаемости этих покровов для воды и растворенных в ней веществ варьирует у разных групп животных. У амфибии кожа обычно влажная, с высокой проницаемостью для кислорода и двуокиси углерода, а также для воды и ионов, которые проходят через нее посредством пассивной диффузии. Компенсация потери электролитов у амфибий идет за счет активного транспорта через кожу внутрь организма солей из окружающей водной среды. У рыб обязательно проницаемыми должны быть жабры, потому что они участвуют в обмене О2 и СО2 между кровью и внешней водной средой. Через жабры осуществляется также активный транспорт солей. У наземных животных (рептилий, птиц, млекопитающих, насекомых) кожа, как правило, намного хуже пропускает жидкость, и в результате потеря воды через кожу протекает обычно медленнее. Те представители данных классов, которые в ходе эволюции вновь стали водными, например прудовые насекомые и морские млекопитающие, сохранили свои внешние покровы тела малопроницаемыми.
Как правило, проницаемость внешних покровов тела для воды у животных с высоким уровнем осморегуляции меньше, чем у тех животных, у которых подобная регуляция развита слабо. У некоторых видов уменьшение проницаемости покровов выглядит как реакция в ответ на осмотическое воздействие среды. У рыб, устойчивых к широкому диапазону осмотических параметров водной среды, адаптация к соленой воде выражается резким падением проницаемости внешних покровов. Как известно, приток крови к жаберному эпителию у некоторых рыб ослабляется при снижении интенсивности дыхания и, наоборот, усиливается, когда потребность в кислороде возрастает. Это сокращает излишнюю циркуляцию крови через жабры, а следовательно, ограничивает ненужный в данном случае осмотический обмен через жаберный эпителий.
4.Питание. Во время питания животные поглощают воду и растворенные в ней вещества. В пище может содержаться избыток как воды, так и солей. Чайки, питаясь прибрежными морскими беспозвоночными, поглощают относительно много соли по сравнению с водой. В связи с этим у них должны быть развиты специальные органы для удаления избытка соли. Пресноводные рыбы и амфибии, напротив, потребляют мало соли по сравнению с тем количеством воды, которое они выпивают, и поэтому им необходимы специальные органы, препятствующие выведению солей из организма.
5.Температура, физическая нагрузка и дыхание. Вода с ее высоким значением теплоты парообразования идеально подходит для удаления из организма тепла путем испарения с поверхности эпителия. В процессе испарения молекулы воды, получившие максимальную энергию, переходят в газообразную фазу, унося с собой тепло. В результате оставшаяся на поверхности эпителия вода охлаждается. У наземных животных особая роль воды в терморегуляции тела создает противоречие и своеобразный компромисс между процессами физиологической адаптации к температуре окружающей среды и воздействием осмотического стресса. Животные пустынь, которые сталкиваются с действием высоких температур и в то же время недостаточным
поступлением в ткани воды, испытывают огромные трудности, потому что им приходится одновременно избегать и перегрева, и потери из организма большого количества жидкости. Интенсивная физическая нагрузка не уменьшает, а, наоборот, способствует росту температуры тела из-за усиления обменных процессов в мышцах, и, следовательно, этот
33
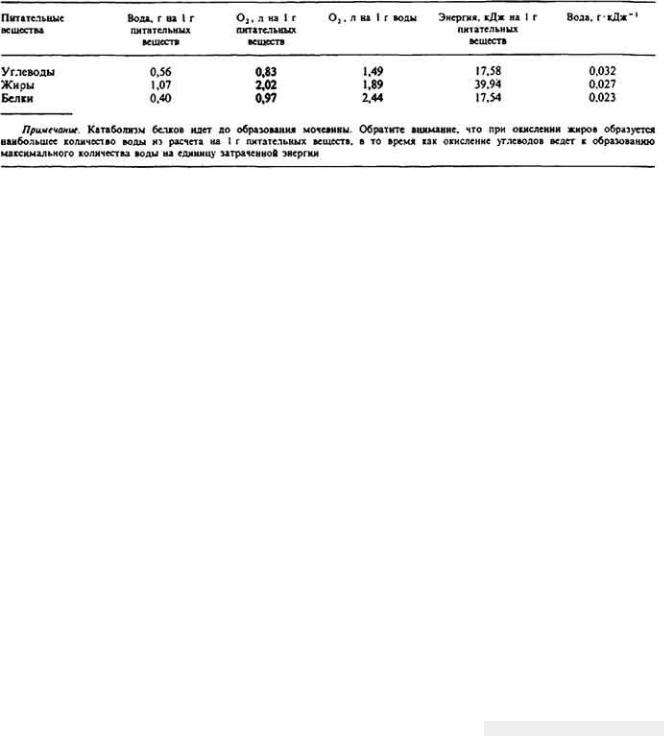
Таблица 12-4. Типичное отношение между образованием метаболической воды, превращением энергии и потреблением О2 при окислении различных питательных веществ до СО2 (Edney, Nagy, 1976)
перегрев* должен быть компенсирован за счет энергичной отдачи тепла в окружающую среду. Наилучший способ такой компенсации - испарительное охлаждение поверхности таких органов, как легкие, воздухоносные пути, язык, или испарение. воды с поверхности кожи. Даже в состоянии покоя (когда отсутствуют движения, за исключением дыхания) механизм работы органов дыхания многих наземных животных таков, что неминуемо ведет к потере воды через газообменные поверхности. Мы вернемся к проблемам регуляции температуры тела в гл. 16.
6. Метаболические факторы. Образуемые в результате пищеварения и обмена веществ конечные продукты, которые организм уже не может использовать, должны удаляться наружу. Так, двуокись углерода диффундирует из организма в окружающую среду через газообменную поверхность. Другой основной конечный продукт клеточного обмена веществ - вода - образуется в довольно малых количествах, и поэтому ее удаление из организма не представляет трудностей (табл. 12-4). На самом же деле эта так называемая метаболическая вода должна сохраняться в тканях у некоторых видов животных. Она служит главным источником влаги для многих обитателей пустыни. Неизбежное образование азотсодержащих конечных продуктов метаболизма, таких как аммиак и мочевина, также создает осмотические проблемы, поскольку для устранения из организма указанных веществ опятьтаки нужна вода.
34
32 :: 33 :: 34 :: Содержание

34 :: 35 :: Содержание
12.1.2. Осморегуляторы и осмоконформеры1
Животные, обладающие способностью поддерживать концентрацию осмотически активных веществ во внеклеточных жидкостях и клетках своего тела на ином уровне, чем во внешней среде, в которой они обитают, были названы осморегуляторами. Животных, которые не могут активно регулировать осмотические свойства жидкостей своего организма, а вместо этого подчиняются осмотическому воздействию окружающей среды, называют осмоконформерами. В табл. 12-2 приведены примеры этих двух крайностей осмотической адаптации. Большинство позвоночных (исключение составляют пластино-жаберные рыбы и миксины) являются истинными осморегуляторами, поддерживающими осмотические показатели жидких сред своего тела в узких пределах. Хотя между видами позвоночных все же существует в осмотическом отношении некоторое различие, кровь позвоночных является гипоосмотической (или, по крайней мере, слегка гиперосмотической, как у акул) в сравнении с морской водой и существенно гиперосмотической в сравнении с пресной. Сказанное справедливо и для тех рыб, которые мигрируют между пресной и морской водой и используют свои эндокринные механизмы, для того чтобы выдержать меняющийся осмотический стресс, обусловленный переходом из одной среды обитания в другую.
У большинства наземных беспозвоночных осмо-регуляция развита хорошо. Конечно, водные беспозвоночные, обитающие в пресной, солоноватой или морской воде, подвергаются неодинаковым осмотическим воздействиям со стороны окружающей среды. Морские беспозвоночные, как правило, находятся в осмотическом равновесии с соленой водой. Из табл. 12-2 видно, что концентрация Na+, К+, Са2+, Mg2+ и Сl- в жидких средах тела морских беспозвоночных близка по значениям к концентрации этих ионов в морской воде, где данные животные обитают. Подобное сходство позволило физиологам, изучающим ткани морских животных, использовать морскую воду в качестве физиологического раствора. Концентрация ионов во внутренних жидких средах различных пресноводных и наземных беспозвоночных отличается от концентрации во внешней среде гораздо сильнее, чем у морских беспозвоночных. Внутренние жидкости у данных животных всегда более разбавлены, чем
34
морская вода, но значительно концентрированнее, чем пресная.
Среди водных беспозвоночных одни являются строгими осморегуляторами
(позвоночные), другие - ограниченными осморегуляторами, а третьи -строгими осмоконформерами. На графике (рис. 12-5) показано, как меняется концентрация осмотически активных веществ во внеклеточных пространствах указанных групп животных в зависимости от соответствующей концентрации в окружающей водной среде. По мере изменения концентрации осмотически

активных веществ в окружающей воде внутренняя концентрация этих веществ у строгих осмо-конформеров меняется аналогичным образом, и ее линия не графике следует параллельно линии, отображающей осмотическое равновесие между организмом и внешней средой. У строгих осморегуляторов, напротив, внутренняя концентрация осмотически активных веществ поддерживается на постоянном уровне в широком диапазоне изменения концентрации этих веществ во внешней среде. На графике это представлено горизонтальной линией, идущей параллельно оси абсцисс. Ограниченные осморегуляторы поддерживают постоянной осмотическую концентрацию своих внутренних жидкостей в ограниченном диапазоне осмотических концентраций внешней среды. За пределами данного диапазона они функционируют как осмоконформеры.
Таким образом, животные, очевидно, преодолевают большие колебания осмотических условий внешней среды двумя способами. Во-первых, они могут вести себя как осмоконформеры, проявляя в высокой степени осмотическую толерантность клеток. Во-вторых, они способны существовать как осморегуляторы и строго поддерживать внеклеточный осмотический гомеостаз в ответ на большие перепады концентрации электролитов во внешней среде. Ткани осморегуляторов обычно не в состоянии противостоять даже более чем мизерным изменениям осмотической концентрации внеклеточной жидкости и поэтому вынуждены всецело зависеть от осмотической регуляции внеклеточных жидкостей. С другой стороны, клетки осморегулирующих животных могут преодолевать действие высокой концентрации осмотически активных веществ в плазме за счет увеличения собственной осмотической концентрации. Один из способов достижения данной цели состоит в увеличении концентрации внутриклеточных органических осмолитиков - молекул, чье присутствие в больших концентрациях повышает осмотическую активность цитоплазмы. Наличие таких молекул уменьшает необходимость поддержания осмотического давления за счет неорганических ионов, которые могли бы вызвать другие осложнения, например ослабить активность ферментов.
Рис. 12-5. Зависимость между осмотической
концентрацией жидких сред организма и окружающей среды для трех классов водных животных.
У некоторых морских позвоночных и беспозвоночных органические осмолитики обнаружены также в крови и тканевых жидкостях. Поэтому внеклеточная осмотическая концентрация у данных животных близка к таковой в морской воде. Наиболее известными примерами указанных осмолитиков

служат мочевина и оксид триметиламина. Оба вещества используются различными морскими пластиножаберными рыбами, примитивной рыбой Latimeria из группы целакантообразных и питающейся крабами лягушкой Rana cancrivora, которая обитает в солоноватых водоемах Юго-Восточной Азии (табл. 12-2).
35
1 Гомойосмотические в пойкилосмотические животные.- Прим. перев.
34 :: 35 :: Содержание