

63 :: 64 :: 65 :: Содержание
12.5.2. Осморегуляторные органы секреторного типа
Мальпигиевы сосуды вместе с задней кишкой образуют у насекомых основной экскреторно-осморегуляторный орган. В общих чертах он (рис. 12-33) состоит из длинных тонких мальпигиевых сосудов, которые впадают в пищеварительный канал в месте соединения средней и задней кишки. Слепые концы мальпигиевых сосудов находятся в гемоцеле (полости тела, содержащей кровь). Секрет, образующийся в сосудах, перетекает в заднюю кишку, где он обезвоживается, а затем поступает в прямую кишку и выводится из организма через анальное отверстие в виде концентрированной мочи. С развитием у насекомых трахейного дыхания, которое рассматривается в гл. 14, значение кровеносной системы в указанном процессе уменьшается. В результате мальпигиевы сосуды не получают артериальную кровь непрерывно под давлением как нефроны млекопитающих. Зато они окружены кровью, давление которой по существу не превышает давление в сосудах. Поскольку у насекомых значительной разницы давлений по обе стороны стенки мальпигиевых сосудов нет, фильтрация не может сыграть у них заметной роли в образовании мочи. Поэтому данный процесс у насекомых должен идти полностью за счет секреции веществ и, возможно, с последующим обратным всасыванием некоторых компонентов секретируемой жидкости; он аналогичен образованию мочи за счет секреции в бесклубочковых почках морских костистых рыб. Серозная поверхность мальпигиева сосуда сплошь покрыта микроворсинками, и клетки, образующие эти сосуды, очень богаты митохондриями (рис. 12-34). Такие особенности структуры органа часто связаны с высокой активностью секреторного эпителия.
Детали процесса образования мочи в мальпигиевых сосудах неодинаковы у разных видов насекомых, но некоторые основные его черты, по-видимому, всегда совпадают. Калий и в меньшей степени натрий активно секретируются в просвет сосуда вместе с конечными продуктами азотистого обмена (мочевой кислотой, аллантионом). Активный перенос К+ в просвет сосуда, по-видимому, служит главной движущей силой процесса образования мочи. При этом большинство остальных веществ
63

Рис. 12-33.
Упрощенная схема выделительной системы насекомого. Первичная моча секретируется в просвет мальпигиевых сосудов, а затем переходит в прямую кишку, где она концентрируется за счет удаления воды. Уменьшение объема мочи в прямой кишке вызвано реабсорбцией воды. Стрелками указан путь циркуляции воды и ионов (см. описание в тексте). На рисунке изображены полько два мальпигиевых сосуда, хотя на самом деле их множество. (Wigglesworth. 1932.)
Рис. 12-34.
Секреторная клетка в стенке мельпигиевого сосуда таракана. Поверхность, обращенная в сторону мочи, увеличена за счет тесно прилегающих друг к другу микроворсинок (некоторые ворсинки утолщены из-за присутствия в них митохондрий). На поверхности клеточной мембраны, обращенной в сторону гемолимфы, имеются беспорядочно расположенные в базолатеральном направлении щели, выполняющие важную роль в осмотическом движении жидкости. (Oschman, Berridge, 1971.)
переходит в просвет сосуда пассивно. Данный вывод был сделан на основании следующих наблюдений:
1.В мальпигиевых сосудах моча изотонична или слегка гипертонична относительно гемолимфы.
2.У всех насекомых моча в сосудах имеет высокую концентрацию К+.
3.Скорость образования мочи в сосудах зависит от концентрации К + в смывающей их жидкости. Чем выше концентрация калия, тем быстрее идет процесс накопления мочи.
4.Образование мочи в мальпигиевом сосуде почти не зависит от концентрации натрия в окружающей выделительный орган жидкости.
Несмотря на то что наиболее важное в осмотическом плане вещество из активно переносимых через мембрану в просвет сосуда, это-калий, есть данные, свидетельствующие о существенной роли активного транспорта в секреции мочевой кислоты и других азотсодержащих конечных продуктов метаболизма.
Первичная моча, образовавшаяся в мальпигиевых сосудах, довольно однообразна по составу у разных видов животных и у каждого вида сохраняет свою изотоничность по отношению к гемолимфе при разных уровнях активности осморегуляторной системы организма. Образовавшаяся в мальпигиевых сосудах моча поступает в заднюю кишку, где ее состав существенно изменяется. Ионы и вода удаляются из мочи в количестве, необходимом для сохранения надлежащего состава гемолимфы. Таким образом, именно задняя кишка определяет состав конечной мочи.
Вода и ионы, извлекаемые из мочи в задней кишке, проходят через тесно контактирующие структуры клеток в просвет мальпигиевых сосудов, т.е. данные вещества накапливаются и рециркулируют в цепи "мальпгиев сосуд-задняя кишка".
Наиболее полное изучение осморегуляторной функции задней кишки было проведено на пустынной саранче Schistocerca. Когда в заднюю кишку этих насекомых вводили раствор, сходный по составу с гемолимфой, то вода, а также ионы калия, натрия и хлора всасывались в гемолимфу, омывающую экскреторный орган. Данные, полученные на основе электрических измерений, позволили сделать предположение об активном переносе ионов через стенку кишки. Задняя кишка саранчи в состоянии осуществлять всасывание большого количества воды, в результате чего моча становится гипертонической, превосходя кровь по осмотической концентрации в 4 раза. Поглощение воды — процесс активный (т.е. сопряженный с гидролизом молекулы АТР, которая служит источником энергии) и поэтому происходит даже против осмотического градиента, который соответствует пятикратному превосходству осмотического давления содержимого прямой кишки над таковым гемолимфы. У "мучного червя" (личинки жука хрущака мучного, Tenebrio) осмотическое отношение моча/кровь может достигать 10, что сравнимо с концентрирующей
64
способностью наиболее эффективного экскреторного органа - почкой млекопитающих. Возможен и альтернативный, чисто физический механизм, который может обеспечить перенос воды через клеточные структуры эпителия без непосредственной связи с высвобождением химической энергии. Например, полагают, что у некоторых видов, таких как Tenebrio, этот перенос воды обусловлен расположением мальпигиевых сосудов перинефридиального пространства и прямой кишки, которые все вместе образуют противоточный концентрирующий механизм (рис. 12 - 35). В соответствии с результатами Филлипса (Phillips, 1970) вода поступает из прямой кишки в мальпигиевы сосуды под действием осмоса, обусловленного появлением градиента концентрации КСl из-за активного переноса этих ионов. Основной поток

веществ в указанных компартмeнтах ориентирован так, чтобы осмотический градиент вдоль экскреторного органа был максимальным, причем абсолютно высокая осмотическая концентрация имела место ближе к анальному концу прямой кишки. Этот градиент позволяет создать концентрацию жидкости, находящейся возле анального конца прямой кишки, в несколько раз превышающую концентрацию гемолимфы. Более того, рециркуляция КСl (рис. 12-33) в мальпигиевых сосудах и задней кишке создает в перинефридиальном пространстве и просвете мальпигиева сосуда осмотическое давление, необходимое для извлечения части воды из прямой кишки. Таким образом, механизм стационарного градиента концентрации, по-видимому, обусловливает в задней кишке насекомых перенос воды вслед за растворенным в ней веществом (К.С1).
Рис. 12-35.
Водно-экстракционный механизм прямой кишки жука мучного хрущака Tenebrio. Основная часть воды и КСl. поступивших в просвет прямой кишки, рециркулирует в мальпигиевых сосудах. (Phillips. 1970.)
Мы очень мало знаем о регулировании осмотического давления у беспозвоночных по принципу обратной связи, однако есть данные о существовании такого регулирования у насекомых. Клоп Rhodnius раздувается после того, как напьется крови из тела своего хозяина - млекопитающего. Но уже через 2 - 3 мин мальпигиевы сосуды увеличивают секрецию жидкости более чем в тысячу раз, обильно продуцируя мочу. Искусственное введение солевого раствора "голодному" насекомому не вызывает такого бурного диуреза. Было также показано, что изолированные мальпигиевы сосуды, погруженные в гемолимфу "голодных" клопов, остаются неактивными. Но если их опустить в гемолимфу клопов Phodnius, которые недавно напились крови, произойдет обильная секреция жидкости. Вещество, стимулирующее процесс секреции в данных сосудах, можно экстрагировать из нервной ткани, содержащей тела или аксоны нейросекреторных клеток, в особенности клеток заднегрудного ганглия. Таким образом, эти клетки, по-видимому, выделяют диуретический гормон в ответ на какое-то вещество, присутствующее в поглощенной крови. Единственным нейрогуморальным агентом, относительно которого установлено, что он стимулирует диуретическое действие нейросекреторных

клеток, является серотонин. Сходные данные, полученные о других видах насекомых, говорят о регулировании секреторной активности мальпигиевых сосудов или реабсорбирующей активности прямой кишки, диуретическими и антидиуретическими гормонами, выделяемыми нервной системой. Удаление переднего нервного ганглия у дождевого червя приводит к задержке воды в организме и одновременному снижению осмотической концентрации плазмы. Введение гомогенизированной ткани головного мозга устраняет эти эффекты, что указывает на гуморальный механизм данного явления.
65
63 :: 64 :: 65 :: Содержание

66 :: Содержание
12.6. Осморегуляция в водной среде
Как само тело водных животных, так и все их газообменные поверхности, погружены в воду. Осмотическая концентрация водной среды может составлять от нескольких миллиосмолей на литр (в пресных озерах) до приблизительно 1000 мос-моль л-1 (в обычной морской воде) и даже выше (в закрытых морях). В природе существуют и всевозможные среды обитания, промежуточные по солености воды между такими крайностями - солоноватые болота, топи и эстуарии. Как правило, жидкие среды организма (т.е. тканевая жидкость и кровь) защищены от влияния экстремальных осмотических условий окружающей среды. Некоторые водные животные, называемые эвригалинными, могут выживать в широком диапазоне солености воды, тогда как другие, называемые стеногалинными, не выдерживают значительных изменений солености воды. Далее рассматривается, каким образом водные животные преодолевают осмотические нагрузки, начиная с пресноводных животных.
l12.6.1. Пресноводные животные
Внутренние жидкости организма пресноводных животных, будь то позвоночные, рыбы, амфибии, рептилии или млекопитающие, обычно гиперосмотичны по сравнению с их водным окружением (табл. 12-2). Осмотическая концентрация крови пресноводных позвоночных находится в диапазоне от 200 до 300 мосмоль-л-1, тогда как вода обычно имеет осмолярность, равную 50 мосмоль л-1. Так как животные гиперосмотичны по отношению к окружающей водной среде, они сталкиваются с двумя типами осморeгуляторных проблем. Во-первых, пресноводные животные подвергаются набуханию из-за поступления воды в их тело по осмотическому градиенту. Вовторых, поскольку окружающая среда содержит мало солей, животные сталкиваются с таким явлением, как постоянная утечка из организма солей. Таким образом, у пресноводных животных должны предотвращаться реальная потеря солей и реальное накопление воды, что достигается у них несколькими способами.
Истинное накопление воды в организме частично предотвращается удалением се избытка, как в том случае, когда почки продуцируют разбавленную мочу. Так, среди близких видов рыб, те, которые живут в пресной воде, выделяют мочу наружу более обильно (т. е. в большом количестве и, следовательно, разбавленную), нежели их морские родичи (рис. 12-36 и 12-37). Моча становится разбавленной, так как нужные для организма соли в большом количестве задерживаются благодаря обратному всасыванию в почечных канальцах из первичной мочи в кровь. И все-таки некоторое количество солей выводится из организма с мочой. Поэтому всегда существует потенциальная опасность постепенного вымывания из организма таких биологически важных солей, как КCl, NaCl, СаСl2 и MgCl2. Потеря солей частично восполняется за счет потребления пищи. У пресноводных организмов важным адаптивным

механизмом восполнения солевых ресурсов служит активный перенос солей из внешней разбавленной среды через эпителий в тканевую жидкость и кровь. Данный активный транспорт осуществляется эпителиальными клетками, содержащимися, например, в коже у земноводных и жабрах у рыб. Не только у рыб, но и у многих водных беспозвоночных жабры являются основным органом осморегуляции, более эффективным в этом отношении, чем почки.
Пресноводные животные обладают удивительной способностью поглощать соли из разбавленной водной среды. Например, пресноводные рыбы с помощью своих жабр могут извлекать Na+ и Cl- из воды, содержащей менее 1 мМ NaCl, в то время как концентрация NaCl в плазме превышает 10 мМ (рис. 12-37). Таким образом, активный перенос NaCl в жабрах идет против более чем стократного концентрационного градиента. У некоторых пресноводных животных, например рыб, рептилий, птиц и млекопитающих, потребление воды и потеря солей сводятся к минимуму за счет низкой проницаемости внешних покровов тела для данных веществ. Как правило, пресноводные животные, кроме рептилий, птиц и млекопитающих, у которых покровы тела относительно непроницаемы, могут воздерживаться от питься пресной воды, уменьшая тем самым потребность организма в выведении избытка влаги наружу.
66
66 :: Содержание

66 :: 67 :: 68 :: 69 :: Содержание
l12.6.2. Морские животные
Обычно внутренние жидкие среды морских беспозвоночных, включая асцидий (примитивных хордовых), близки к морской воде как по осмолярности, так и по индивидуальным концентрациям в плазме основных неорганических солей (табл. 12-2). Следовательно, для регулирования осмолярности жидких сред своего организма такие животные не нуждаются в больших затратах энергии. Среди позвоночных особое место занимают миксины, у которых плазма изоосмотична по отношению к внешней среде. Однако они отличаются от большинства морских беспозвоночных способностью регулировать в своем теле
концентрацию отдельных ионов. В частности, концентрация Са2+, Mg2+ и SO 2-
4
в теле миксин поддерживается на значительно более низком уровне, чем в морской воде, а концентрация Na + и Сl-, напротив, на более высоком. Поскольку у позвоночных животных различные функции таких
66

Рис. 12-36.
Обмен воды и солей у некоторых видов позвоночных. Обозначен только активный обмен. Пассивная потеря воды через кожу, легкие и пищеварительный тракт не обозначена.
возбудимых тканей, как нервная и мышечная, особенно чувствительны к концентрации Са2+ и Mg2+, механизмы, регулирующие концентрацию этих двухвалентных катионов, развивались у них в тесной связи с особенностями нейромышечного аппарата.
Плазма крови хрящевых рыб-акул, скатов, а также примитивной кистеперой рыбы Latimeria, как и у миксин, изоосмотична по отношению к морской воде. Однако перечисленные животные отличаются гораздо более низкой концентрацией в организме электролитов (например, неорганических ионов) по
67
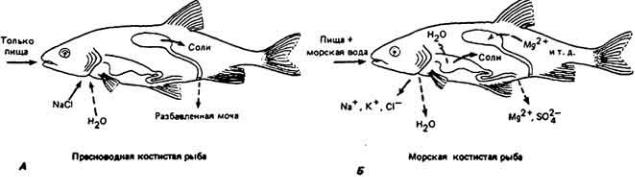
Рис. 12-37.
Обмен соли и воды у пресноводных (А) и морских (Б) костистых рыб. Сплошными стрелками показаны активные процессы, пунктиром - пассивные. Обратите внимание на роль жабр в активном транспорте соли у тех и других рыб. (Prosser, 1973.)
сравнению с концентрацией органических осмолитиковмочевины и оксида триметиламина. У пластиножаберных и кистеперых избыток неорганических электролитов, например NaCl, выводится через почки и специальный экскреторный орган - ректальную железу, находящуюся в конце пищеварительного тракта.
У морских костистых рыб, как и у большинства высших позвоночных, внутренние жидкие среды гипотоничны по отношению к морской воде. Поэтому у таких рыб наблюдают склонность к потере воды в окружающую среду, особенно через жаберный эпителий. Чтобы восполнить потерянный объем воды, они пьют соленую воду (рис. 12-36 и 12-37). От 70 до 80% выпитой морской воды вместе с большей частью содержащихся в ней NaCl и KCl поступает в кровоток благодаря процессу абсорбции через эпителий кишечника.
Большинство двухвалентных ионов - Са2+, Mg2+ и SO 2- задерживается в кишке и
4
затем выбрасывается наружу через анальное отверстие. Избыток соли, абсорбированной вместе с водой, в дальнейшем уходит из крови в морскую воду за счет активного переноса Na+, Cl- и, возможно, К+ через жаберный эпителий, а также путем секреции солей двухвалентных металлов почками (рис. 12-37). Моча в этом случае изотонична по отношению к крови, но богата теми солями,
которые не секретируются жабрами, особенно Mg2+ , Са2+ и СО 2- . Общий итог
4
совместной работы жабр и почек у морских костистых рыб - истинное накопление в организмe воды, гипотоничной по отношению и к выпитой морской воде, и к моче. Пользуясь описанным механизмом, некоторые виды костистых рыб, например лососевые северо-западной части Тихого океана, могут в той или иной степени поддерживать постоянной осмотическую концентрацию плазмы, несмотря на миграцию из моря в реки и обратно.
Подобно морским костистым рыбам морские рептилии (например, игуаны, морские черепахи из эстуариев рек, крокодилы, морские змеи), а также морские птицы не могут выделять мочу, которая была бы намного осмотичнее их внутренних жидких сред (рис. 12-36). Но природа наделила их органом, расположенным на голове и приспособленным для секреции солей в виде
сильно гиперосмотической жидкости. Это так называемые солевые железы, о которых мы уже говорили в разд. 12.4.2. У птиц они обычно находятся на клюве, под глазами, а у ящериц возле носа или глаз. Недавно солевые железы обнаружили в языке крокодилов, живущих в солоноватых водах (существование у этих рептилий отличных от почек средств выведения солей из организма предполагалось давно). Хотя почки рептилий и птиц не могут продуцировать сильно гипертоническую мочу, у морских рептилий и птиц достаточное количество соли выделяется через солевые железы. Поэтому данные животные могут пить соленую воду, несмотря на то что выделяют мочу менее концентрированную, чем морская вода (рис. 2-38). Солевые железы, подобно жабрам морских костистых рыб, компенсируют неспособность почек рептилий и птиц выделять сильно гипертоническую относительно жидких сред организма мочу.
Морские млекопитающие, к которым относятся ластоногие (например, морские львы, тюлени) и китообразные (дельфины, киты), не имеют отличных от почек солевыделительных органов, подобных солевым железам птиц и рептилий или жабрам рыб. Однако природа наделила их, как впрочем в всех остальных млекопитающих, высокоэффективными почками, способными выделять сильно гипертоническую мочу. И тем не менее морские млекопитающие не могут пить соленую воду. Они довольствуются только той влагой, которая
68
Рис. 12-38.
А. Приспособленность чайки к поддержанию водно-солевого баланса. Птица

может пить морскую воду, потому что через солевые железы она выводит из своего организма 80% поглощенной с питьем соли и только 50% воды. В итоге чайка выделяет гипотоническую мочу без обезвоживания организма. Б. Человек и большинство наземных млекопитающих не в состоянии употреблять для питья морскую воду, поскольку их моча не достигает концентрации, достаточной для того чтобы при этом организм сохранял воду и одновременно избавлялся от поглощенной соли. (К. Schmidt-Nielsen. Sail Gland, © /959. Scientific American, Inc.
Все авторские права сохранены.)
содержится в их пище. Образно говоря, они как бы пользуются осморегуляторными приспособлениями своих жертв, в значительной мере избегая той осмотической работы, которая выпадает на долю костистых рыб, рептилий и птиц, пьющих морскую воду (рис. 12-36). Как уже упоминалось, у некоторых пустынных, а также и морских млекопитающих существует еще один источник поступления воды в организм. Речь идет о метаболической воде, образующейся в ходе обменных превращений питательных веществ. В результате этих превращений атомы водорода реагируют с атомами кислорода с образованием молекул воды (см. рис. 3-30, табл. 12-4). Человек, как и другие млекопитающие, не приспособен пить морскую воду. Его почки могут удалить из кровотока примерно до 6 г Na+ на каждый литр образующейся мочи. Морская же вода содержит Na+ в концентрации около 12 гл -1. Поэтому употребление морской воды приведет к накоплению солей в организме, если он не получит физиологически эквивалентного количества пресной воды (рис. 12-38). Иначе говоря, чтобы удалить соль, поступившую в организм вместе с определенным объемом выпитой морской воды, почки человека должны выделить еще больший объем воды. Это непременно приведет к быстрому обезвоживанию организма.
Обобщая все сказанное, подчеркнем, что пресноводные животные склонны потреблять воду пассивно, а удалять ее активным путем за счет осмотической функции почек (позвоночные) или почкообразных нефридиев (беспозвоночные). Они отдают соли в окружающую разбавленную водную среду, восполняя их убыль посредством активной абсорбции ионов из этой среды через кожу, жабры или другие виды эпителия, осуществляющего активный транспорт веществ. С другой стороны, у морских позвоночных осмотическая потеря воды происходит через жабры или покровы тела (если они проницаемы). Чтобы возместить эту потерю, животные пьют морскую воду и активно выводят обратно в окружающую среду избыток солей, поглощенных во время питья. Этот процесс идет благодаря активному переносу ионов через эпителий отличных от почек осморегулирующих органов-жабр и солевых желез. Те морские млекопитающие, у которых нет солевых желез или сходных приспособлении, соленой воды не пьют и довольствуются влагой, которая содержится в съедаемой пище. Их осмотический баланс поддерживается прежде всего за счет работы почек. Морские беспозвоночные животные, как правило, осмоконформаторы.
69
66 :: 67 :: 68 :: 69 :: Содержание