


15 :: 16 :: Содержание
11.2.2. Тонкая структура двигательной органеллы
Основными структурами в ресничке и жгутике являются .микротрубочки. Они тянутся от одного конца двигательной органеллы до другого. Иерархические уровни организации микротрубочек показаны на рис. 11-12. Стержень типичной реснички или жгутика образован девятью парами микротрубочек, расположенных по кольцу вокруг одной центральной пары. Эти девять дублетов и две центральные микротрубочки вместе со вспомогательными структурами формируют аксонему. которая полностью изолирована от внешней среды за счет выпячивания клеточной мембраны.
Микротрубочки аксонемы состоят из тубулиновых глобулярных гетеродимеров, каждый из которых включает в себя один α- и один β-мономер. В состав каждой микротрубочки центральной пары входит 13 протофиламентов, образованных вытянутой цепочкой димеров тубулина. Микротрубочки девяти наружных дублетов устроены сходным образом (рис. 11-13), но здесь протофиламенты, собранные из димеров тубулина, формируют трубчатую структуру как с полным набором протофиламентов (трубочка1 А), так и с неполным набором (трубочка В, прикрепленная к трубочке А).
Если рассматривать поперечное сечение аксонемы по направлению от основания жгутика к его концу, можно увидеть, что каждая трубочка А имеет две боковые ручки, направленные в сторону трубочки В следующего (по часовой стрелке) дублета. Белок, из которого построены эти так называемые динеиновые ручки (они повторяются через определенные интервалы по всей длине трубочки А), проявляет Mg2+-зависимую АТРазную активность. До сих пор принято считать, что динеин -единственный компонент аксонемы, обладающий активностью АТРазы, хотя не исключено присутствие в аксонеме и других АТРаз.
Каждый периферический дублет микротрубочек несет также целую цепочку радиальных спиц. У некоторых жгутиков спицы располагаются парами по всей длине дуплета, как показано на рис. 11-13. Каждая радиальная спица соединена с соответствующим ей выступом на центральной капсуле1 аксонемы. У основания реснички или жгутика находится базальное тельце (его называют также кинетосомой). Оно гомологично центриолям - клеточным органеллам, из которых развиваются микротрубочки, например нити веретена в делящихся клетках. Базальное тельце имеет вид короткого толстого
15
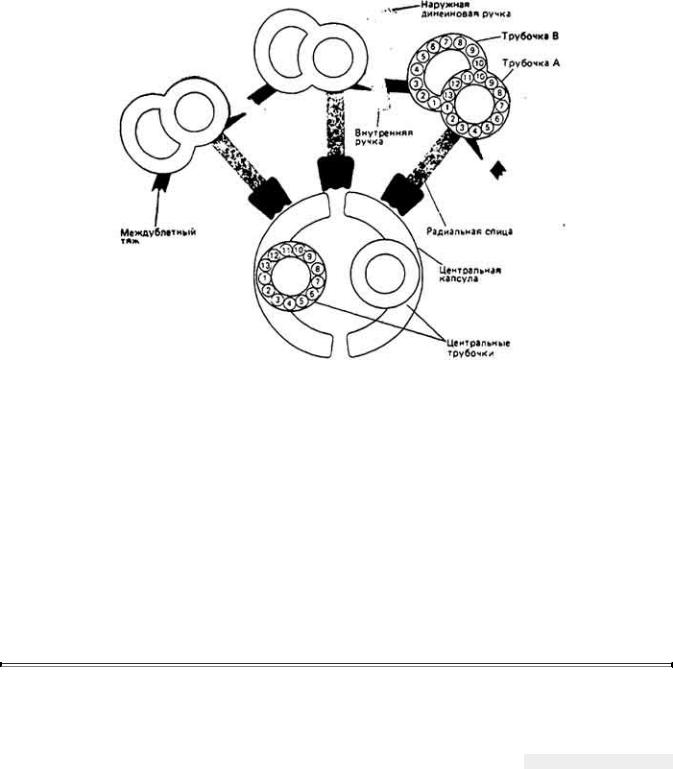
Рис. 11-13. Организация дублетов микротрубочек и вспомогательных структур. Вид а разрезе дан в направлении от основания к концу жгутика. Динеиновые ручки направлены от трубочки А к трубочке В соседнего дублета. Это изображение структуры аксонемы в виде очень простой схемы основано на данных электронно-микроскопического исследования ресничек двустворчатого моллюска Elliptio. (Warner, Satir, 1974.)
цилиндра, наружная стенка которого состоит из 9 триплетов микротрубочек.
Хотя микротубулярная организация жгутиков и ресничек в подавляющем числе случаев относится к типу "9 + 2", все же обнаружены отклонения от этой основной схемы. В зависимости от вида организма данная структура может описываться формулой 3 + 0,6 + 0,9 + 0,9 +1,9 + 3 или 9 + 7. В некоторых случаях периферический ряд микротрубочек включает в себя более 9 дублетов.
16
1Синоним: субфибрилла.- Прим. перев.
1Белковая оболочка, покрывающая центральную пару мигротрубочек.- Прим. перев.
15 :: 16 :: Содержание

16 :: 17 :: Содержание
11.2.3. Химические превращения в ресничках н жгутиках
Определение местонахождения в аксонеме различных белковых компонентов и АТРазы стало возможным благодаря методу химического фракционирования, примененному Гиббонсом (Jan. R. Gibbons) в 1965 г. при изучении жгутиков Tetrahymena. После отделения ресничек от клетки их прежде всего "обрабатывают детергентом для освобождения от клеточной мембраны и аморфного вещества матрикса органеллы. Затем оставшуюся аксонему диализуют в растворе низкой ионной силы, содержащем ЭДТА-вещество, образующее хелаты с Са2+ и Mg2+. Эта обработка позволяет удалить обе центральные микротрубочки и динеиновые ручки всех девяти наружных (периферических) дублетов. Сама же кольцевая структура из девяти наружных дублетов при этом не затрагивается. Ультрацентрифугированием диализата (экстракта, полученного при диализе) выделяют две основные белковые фракции. Та фракция, которая оседает медленно, содержит белки двух центральных фибрилл. Другая фракция состоит из мономерного линейна, обладающего АТРазной активностью, и полимеров разной длины, которые быстрее оседают при центрифугировании. Гиббоне показал, что для проявления ферментативной активности линейна, в результате которой происходит высвобождение неорганического фосфата из АТР (см. рис. 3-9), необходимо присутствие двухвалентного иона, такого как Mg2+.
Если очищенную фракцию линейна смешать с подвергнутой экстракции аксонемой, динеин вновь прикрепляется к трубочкам А наружных (периферических) дублетов, воссоздавая ранее экстрагированные ручки. Учитывая характер расположения динеяновых ручек в аксонеме и их АТРазную активность, можно предположить, что в двигательной органелле они выполняют такую же функцию, как поперечные мостики в поперечнополосатой мышце. Ниже мы опишем, каким образом динеиновые
16
ручки используют энергию, освобождаемую при гидролизе АТР, и генерируют силы скольжения между соседними трубочками.
17
16 :: 17 :: Содержание
17 :: 18 :: Содержание
11.3. Механизмы биения жгутиков
В отличие от сокращения мышечного волокна, которое сводится к укорочению в одном измерении, активные движения ресничек или жгутиков могут осуществляться и в одной плоскости, и в трехмерном пространстве. Это намного усложняет изучение механизма их движения по сравнению с мышечным. К тому же жгутики и реснички очень малы и к ним нелегко прикрепить механический датчик для прямого измерения развиваемого механического напряжения. Поэтому у нас до сих пор нет надежной экспериментально обоснованной теории механизма ресничного и жгутикового движения. Тем не менее последние данные дают основание полагать, что в основе этого движения лежит механизм скольжения трубочек.
11.3.1. Гипотеза скольжения трубочек
Есть два возможных варианта участия кольцевой структуры из девяти дублетов микротрубочек в процессе изгибания аксонемы. 1. Дублеты микротрубочек, расположенные ближе к внутренней линии изгиба, сокращаются, как показано на рис. 11-14, А (или, что то же самое, растягиваются микротрубочки, расположенные ближе к внешней линии изгиба). 2. Дублеты, находящиеся ближе к внутренней линии изгиба, скользят относительно других; при этом длина всех дублетов остается постоянной (рис. 11-14, Б). В настоящее время считают, что в основе подвижности ресничек и жгутиков лежит второй из указанных механизмов. Ниже приводится экспериментальное обоснование этого заключения (Peter Satir, 1973).
Удалось показать, что во время сгибания стержня реснички микротрубочки аксонемы скользят друг относительно друга. Тщательное электронномикроскопическое исследование окончаний девяти периферических дублетов автор провел на препаратах ресничек, мгновенно фиксированных тетраоксидом осмия в разных фазах цикла биения. Оказалось, что микротрубочки, находящиеся на внутренней линии изгиба реснички, всегда выступают несколько дальше, чем находящиеся на наружной линии, и это не зависит от фазы колебательного движения, в которой ресничка была зафиксирована (рис. 11-15). Именно такой выступ у конца аксонемы и следовало бы ожидать, если бы микротрубочки во время скольжения друг относительно друга сохраняли свою длину постоянной. Однако сам по себе этот факт не позволяет нам сделать выбор между следующими двумя возможностями: 1) в данном эксперименте наблюдается активное скольжение микротрубочек, которое и вызывает изгибание стержня реснички; 2) здесь имеет место пассивное скольжение, которое осуществляется вследствие изгибания, вызванного механизмом, отличным от активного скольжения.

Рис. 11-14. Два гипотетических механизма активных изгибов
ресничек. А. При сокращении периферических дублетов, расположенных на одной стороне аксонемы. тело органеллы должно изогнуться в ту же сторону. Принцип работы такого механизма совпадает с процессом, происходящим в биметалической пластинке когда изменение температуры вызывает у металлов разное по силе укорочение. Однако данные, подтверждающие существование подобного дифференцированного сокращения в аксонеме. отсутствуют. Б. Если дублеты на одной стороне аксонемы. будучи закреплены в области основания, находятся в состоянии активного скольжения относительно тех. которые расположены на другой стороне, тогда аксонема должна будет согнуться в сторону дублетов, стремящихся скользить в направлении, противоположном основанию органеллы. Повидимому, реснички именно так и сгибаются. (Brokaw, 1968.)
Для активного скольжения нужны физические механизмы, с помощью которых можно развить силу между примыкающими (соседними) дублетами микротрубочек или между дублетами и центральной капсулой аксонемы. В мышцах подобную силу создают поперечные мостики миозина, взаимодействующие с актиновыми филаментами. Напомним, что, в девяти периферических дублетах аксонемы содержатся динеиновые ручки, выступающие из трубочек А в сторону трубочек В. Кроме того, есть еще и радиальные спицы, которые тянутся от трубочек А внутрь к центральной капсуле (рис. 11-13). Поскольку динеиновые ручки проявляют АТРазную активность, вполне вероятно, что они действительно участвуют в превращении химической энергии в механическую.
В начале 1970-х годов было сделано важное открытие - показали способность периферических
17

Рис. 11-15. Электронно-микроскопические данные в пользу гипотезы скользящих трубочек, объясняющей механизм движения ресничек (жгутиков). Кончики ресничек иccледовали в электронный микроскоп после того как движущиеся органеллы были мгновенно зафиксированы с помощью тетраоксида осмия. Установлено, что периферические дублемы. находящиеся на внутренней стороне изгиба, выступают по направлению верхушке реснички дальше, чем остальные, независимо от того, в какой фазе цикла биения ресничка была остановлена. (Satir. 1973.)
дублетов активно скользить друг относительно друга, используя в качестве источника энергии АТР. На первом этапе исследований с целью удаления поверхностной мембраны и растворимых компонентов стержня жгутики спермиев морского ежа обрабатывали детергентом тритон Х-100 (см. дополнение 10-1). Затем экстрагированные аксонсмы на короткое время подвергали действию фермента трипсина, который разрушает определенные пептидные связи. Обработка трипсином так изменяла структуру аксонемы, что последующее воздействие АТР и Mg2+ вызывало сначала ее удлинение, а потом распад на отдельные трубочки или группы трубочек. С помощью микроскопии в темном поле удалось установить, что удлинение аксонемы происходит в результате продольного скольжения девяти периферических дублетов друг относительно друга (рис. 11-13). При этом общая длина взаимно перекрывающихся дублетов могла превысить длину отдельных микротрубочек в 5 раз. Полученные результаты можно объяснить следующим образом. В живой ресничке за счет энергии АТР происходит скольжение между динеиновыми ручками и трубочкой В следующего дублета. Этому активному движению противодействует некий упругий ограничитель, роль которого, вероятно, выполняют междублетные мостики, изображенные на рис. 11-13. Вызванное таким образом ограниченное скольжение микротрубочек собственно и приводит к изгибу жгутика (реснички). Если же определенные структуры аксонемы разрушены трипсином, то добавление Mg2+ и АТР вызывает уже ничем не сдерживаемое скольжение микротрубочек. Оно-то и приводит к нарастающему удлинению общей структуры, потому что первый дублет скользит относительно второго, второй в свою очередь относительно третьего и так далее (рис. 11-16, Б, В). Для обеспечения активного скольжения нужны такие же источники энергии и ионы, как и для реактивации биения в экстрагированных препаратах жгутика (реснички).

В последующих электронно-микроскопических исследованиях на ресничках Tetrahymena было выявлено, что соседние дублеты движутся друг относительно друга таким образом, что трубочка А, несущая динеиновые ручки, скользит в сторону основания трубочки В, с которой она взаимодействует. Следовательно, динеиновые ручки, по-видимому, развивают усилие за счет активных поворотных движений, которые осуществляются по направлению от основания реснички к ее концу.
Итак, движение ресничек и жгутиков скорее всего вызывается механизмом скольжения трубочек, принципиально схожим с механизмом скольжения нитей в мышцах. Остается нерешенным весьма важный вопрос о том, как регулируется процесс скольжения девяти радиально расположенных дублетов, приводящий к сложным движениям жгутиков и ресничек.
18
17 :: 18 :: Содержание