


52 :: 53 :: 54 :: 55 :: Содержание
12.3.8 Механизмы регуляции почечной функции
Животное может испытывать осмотический стресс по разным причинам: из-за колебаний температуры и солености окружающей среды, а также при потреблении пищи и воды. Сдвиги в осмотическом состоянии жидких сред организма сводятся до минимума работой механизмов с обратной связью, которые изменяют активность саморегулирующих органов так, чтобы сохранить внутренний статус-кво. Почечные механизмы регуляции по принципу обратной связи могут быть эндокринными, нервными или нейро-эндокринными. Существует несколько способов регуляции образования мочи в ответ на осмотический и другие сигналы. Она включает управление: 1) скоростью клубочковой фильтрации; 2) скоростью абсорбции солей из просвета почечного канальца и 3) скоростью осмотического удаления воды из первичной мочи.
12.3.8.1. Регуляция скорости клубочковой фильтрации (СКФ)
Поскольку количество соли и воды, проходящее через боуменову капсулу, в 100 раз превышает содержание этих веществ в конечой моче, значительные колебания СКФ привели бы к разрушению водно-солевого баланса, если бы механизмы реабсорбции и секреции не осуществляли удивительно эффективную коррекцию. В почках птиц и млекопитающих кровоток сохраняется относительно постоянным, несмотря на большие колебания в величине системного давления крови. Такая независимость почечного кровотока имеет важное значение для тонкой регуляции скорости и давления крови, поступающей в клубочковый аппарат (эта регуляция играет главную роль в установлении величины СКФ). Изменение СКФ, особенно у низших позвоночных, таких как рыбы и земноводные, служит одним из приемов, с помощью которого почечная система преодолевает осмотический стресс. Так, у форели в пресной воде СКФ высока, но снижается примерно на 90%, когда форель переходит в морскую среду, где рыбе необходимо сохранить в организме внутреннею воду.
Механизмы регуляции СКФ у млекопитающих сложны. Большое влияние на скорость фильтрации оказывает уровень перфузии клубочковых капилляров. В свою очередь этот уровень сильно зависит от того, насколько сужаются почечные артериолы. Последние контролируются как нервной (симпатическими нервами почек), так и эндокринной (почечными гормонами) системой, о чем см. ниже.
Работа почек регулируется симпатической нервной системой, которая иннервирует юкстагломеру-лярный комплекс (ЮГК), изображенный на рис. 1222 и 12-23. ЮГК состоит из плотного пятна
52
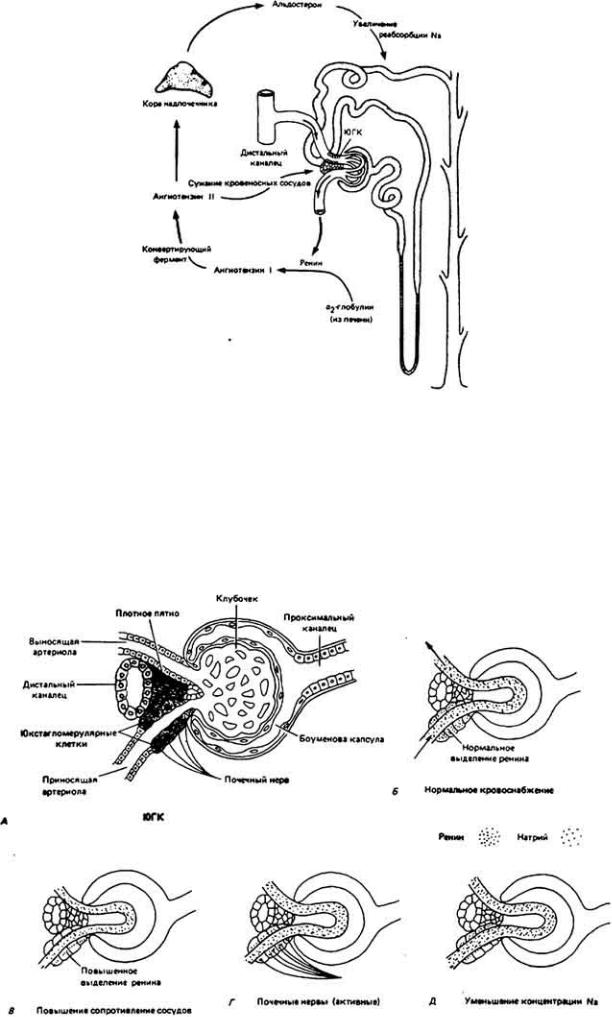
Рис. 12-22.
Петли ренин-ангиотензиновой обратной связи. Юкстагломерулярный комгиекс (ЮГК) состоит из ренинсекретирующих клеток, расположенных преимущественно в стенке приносящей артериолы, и осмочувствительных клеток в плотно прижатой стенке дистального участка почечного канальца. Ренин, высвобождение которого происходит, например, при сужении приносящей
артериолы (из-за снижения давления крови в сосуде или концентрации Na+ в дистальном канальце), приводит к увеличению титра ангиотензина II и
альдостерона. Последний стимулирует активный транспорт Na+ из почечного канальца.
Рис. 12-23.
Юкстагломерулярный комплекс. Выступая в качестве источника ренина. ЮГК имеет два вида реиепторной ткани: сосудистые рецепторы приносящей артериолы. чувствительные к растяжению стенки сосуда, и плотное пятно, реагирующее на концентрацию натрия. Скорость выделения ренина регулируется также почечным нервом. Общая схема действия /ренина показана на рис. 12-22. (J. О. Davis. Control of Renin Release. Hospital Practice 9 (4), 1974; данный рисунок,
помещенный там на стр. 57. взят у Alan Iselin.)
53
(macula densa), образованного примыкающей к клубочку висканальцевой тканью. В него также входят специализированные рецепторные и секреторные клетки, которые окружают участок приносящей артериолы, вплотную примыкающий к дистальному участку почечного канальца. Активация симпатических нервов, иннервирующих ЮГК, приводит к высвобождению из секреторных клеток протеолитического фермента ренина (секреторные клетки расположены в стенке, приносящей артериолы. по которой кровь поступает в клубочковые капилляры боуменовой капсулы). Ренин высвобождается из этих клеток также в ответ на падение концентрации Na+ в плазме или в ответ на такие локальные стимулы, как спадение стенок артериол по причине уменьшения давления крови и концентрации циркулирующих катехоламинов. Считают, что клетки плотного пятна, близко расположенные к ренинсекретирующим клеткам артериолы, должны быть чувствительными к концентрации солей.
Как показано на рис. 12-22, ренин высвобождается клетками ЮГК в просвет приносящей артериолы. Там он действует на α2-глобулин-гликопротеин, синтезируемый в печени и циркулирующий в плазме крови. Ренин расщепляет в молекуле глобулина связь между двумя остатками лейцина, освобождая декапептид (пептид, состоящий из 10 остатков аминокислот) - ангиотензин I. З а т е м конвертирующий фермент отделяет от декапептида еще два аминокислотных остатка, превращая его в октапептид- ангиотензин II. Полученный гормон обладает множественным действием, в частности, способствует общей вазоконстрикции (в том числе сужению ортериол), в результате чего увеличивается кровяное давление, а следовательно, усиливаются как почечный кровоток, так и клубочковая фильтрация.
12.3.8.2. Регуляция канальцевой реабсорбции Na+
Еще одно важное действие ангиотензина II заключается в том, что он стимулирует выделение из коры надпочечников стероидного гормона альдостерона (см. табл. 9-13). Это один из так называемых минералокортикоидов, имеющих первостепенное значение для задержки в тканях солей. Альдестерон действует на дистальный каналец нефрона, повышая скорость реабсорбции натрия, и в конечном счете осмотическую реабсорбцию воды. Он усиливает абсорбцию Na+ из почечного канальца путем стимуляции активного переноса К+ и H+ из плазмы в клубочковый фильтрат. Увеличение переноса Na+ происходит скорее не по причине непосредственного действия натриевого насоса, а из-за индуцированного альдостероном повышения
проницаемости для Na+ клеточной мембраны, расположенной на слизистой стороне канальцевого эпителия (т. е. на клеточной стороне, обращенной в просвет канальца). В результате подобного возрастания проницаемости для натрия канальцевой мембраны возникает усиленный приток ионов Na+ из просвета канальца в эпителиальные клетки (за ними следуют ионы С1-). Подъем концентрации внутриклеточного Na+ обеспечивает насос, находящийся на серозной стороне (т.е. стороне, обращенной к кровеносным капиллярам) клеток с повышенной концентрацией Na+. Таким способом увеличивается скорость переноса Na+ из почечного канальца через слой эпителиальных клеток в тканевую жидкость и кровь1.
Итак, основной эффект ренина, выделяемого в просвет приносящей артeриолы,- усиление реабсорбции NaCl из клубочкового фильтрата. Этот процесс имеет явно регуляторный характер, направленный на компенсацию NaCl, в случае любого уменьшения объема плазмы или концентрации Na* (вспомним, что ренин высвобождается в ответ на падение уровня Na+ в плазме или сужение просвета артериол).
12.3.8.3. Регулирование реабсорбции воды
Канальцевая жидкость, продвигаясь по собирательному протоку в глубокие гиперосмотические области мозгового вещества, теряет воду (по закону осмоса) и концентрируется (рис. 12-21). При таком процессе концентрирования жидкости должно регулироваться количество воды, выводимой из организма вместе с мочой. Скорость, с которой вода за счет осмоса переходит из просвета собирательной трубочки в тканевую жидкость, зависит от проницаемости для воды эпителия, образующего стенки протока. Эта проницаемость (и, следовательно, количество воды, покидающее организм животного с мочой) регулируется антидиуретическим гормоном (АДГ), который секретируется нейрогипофизом (задней дблей гипофиза). Чем выше уровень АДГ в крови, тем более проницаема для воды эпителиальная стенка собирательной трубочки и тем большее количество воды извлекается из мочи по мере движения последней подрубочке к почечной лоханке. Влияние АДГ на реабсорбцию воды из собирательной трубочки показано на рис. 12-24. Поскольку осмотические концентрации в трубочке и тканевой жидкости в эксперименте поддерживали постоянными, то наблюдаемое повышение скорости реабсорбции должно быть следствием увеличенной под влиянием гормона проницаемости стенки трубочки для воды.
54

Рис. 12-24.
Влияние АДГ на осмотическую реабсорбцию воды из собирательной трубочки в тканевую жидкость. Осмотическая концентрация жидкости, проходящей по
трубочке, составляет 125 мосмоль-л01. осмолярность жидкости в кювете - 290
мосмоль-л-1. Период добавления гормона отмечен на графике чертой (Grantham. 1971.)
Уровень АДГ в крови зависит от осмотического давления плазмы. На возрастание последнего чувствительные к данному фактору нейроны, расположенные в гипоталамусе, реагируют повышением частоты импульсации. Это нейросекреторные клетки, аксоны которых достигают нейрогипофиза. При усилении активности данных клеток возрастает скорость высвобождения АДГ в кровоток из окончаний их аксонов и таким образом повышается уровень АДГ в крови. Регуляторный эффект данного нейро-эндокринного механизма изображен на рис. 12-25. Например, если осмотическая концентрация крови возрастает из-за обезвоживания организма, то нейросекреторные нейроны усиливают свою активность, увеличивается секреция АДГ, повышается проницаемость стенок собирательных трубочек и в итоге возрастает скорость удаления воды из мочи за счет осмоса. Описанный процесс приводит к экскреции более концентрированной мочи и сохранению воды в организме. Клетки гипоталамуса, продуцирующие и выделяющие АДГ, получают тормозные сигналы от рецепторов, реагирующих на рост кровяного давления. Эти рецепторы расположены в разных отделах кровеносной системы, но в основном их можно встретить в левом предсердии. Любая причина, ведущая к росту артериального давления (например, увеличение объема крови при употреблении большого количества воды), будет угнетать клетки гипоталамуса, выделяющие АДГ, и, следовательно, увеличивать потерю воды из организма с мочой.
Употребление человеком алкоголя угнетает выделение АДГ, приводя к обильному мочевыделению и росту осмотической концентрации плазмы за пределы нормального уровня. Создается известное обезвоживание организма, которое является одной из причин неприятных ощущении после похмелья.
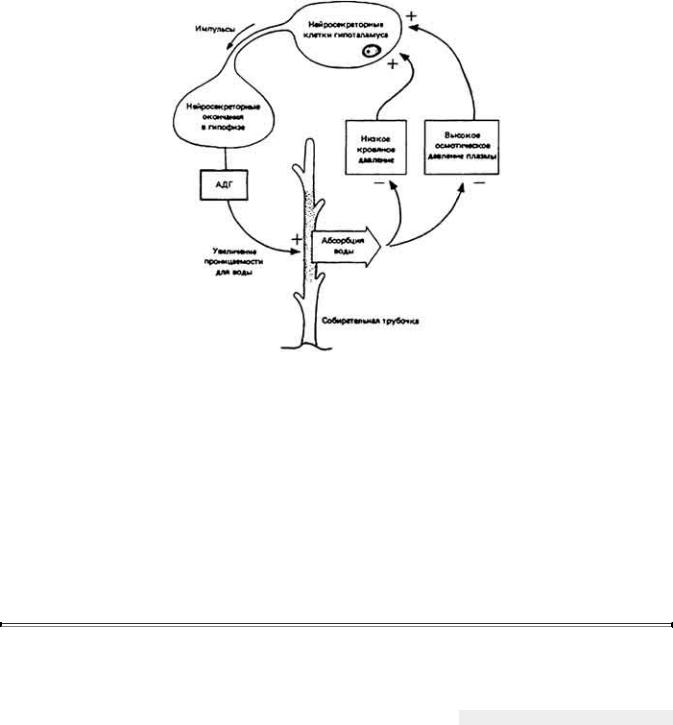
Рис. 12-25.
Регуляция осмотической концентрации крови по принципу обратной связи за счет действия АДГ на собирательную трубочку. АДГ усиливает проницаемость для воды участка трубочки, выделенного точками, в результате чего повышается скорость осмотического удаления воды из мочи. Повышенная реабсорбция воды противодействует усилению состояния, которое стимулирует секрецию АДГ.
АДГ (у млекопитающих) и родственные пептиды (у других классов животных) действуют не только на почки. Аппликация этих гормонов к коже лягушки и мочевому пузырю жабы также повышает проницаемость эпителия данных анатомических структур для воды.
55
1 В некоторых других видах транспортного эпителия, например стенке мочевого пузыря и покровах тела амфибий (см. гл. 4), альдостерон действует таким же образом. У млекопитающих данный гормон способствует реабсорбции Na+ также в слюнных и потовых железах и ободочной
52 :: 53 :: 54 :: 55 :: Содержание