

59 :: 60 :: 61 :: Содержание
12.4.2. Секреция веществ солевыми железами у птиц и рептилий, не связанная с почками
В 1957 г. Шмидт-Нильсен (Knut Schmidt-Nielsen) с соавторами, исследуя, каким способом морские птицы поддерживают свой осмотический баланс без поступления извне пресной воды, нашли у них в носу солевые железы, выделяющие гипертонический раствор NaCl. В этих ранних исследованиях было установлено, что если бакланам или чайкам вводить морскую воду внутривенно или через желудок, то повышение концентрации соли в плазме сопровождается последующим длительным выделением через клюв жидкости, осмотическая концентрация которой в 2-3 раза выше, чем у крови. Спустя некоторые время солевые железы были описаны у многих видов птиц и рептилий, особенно у тех, которые подвергаются осмотическому стрессу, обитая в морях или в пустыне. Эти виды включают в себя почти всех морских птиц, страусов, морскую игуану, морских змей, морских черепах, а также многих наземных пресмыкающихся. У крокодилов подобные железы находятся в языке. Солевые железы птиц и некоторых пресмыкающихся заполняют небольшие углубления в костях черепа (в надглазничной области) (рис. 12-28). У птиц солевые железы состоят из многочисленных долей диаметром около I мм. Каждая долька выводит свой секрет через ветвящиеся секреторные трубки и центральный канал в проток, который в свою очередь проходит по клюву и оканчивается в ноздрях. Активное выделение соли происходит через эпителий ветвящихся трубок, состоящих из особых секреторных клеток. По всей базальной стороне мембраны этих клеток имеются глубокие складки. Кроме того, клетки буквально набиты митохондриями. Как и во всяком другом эпителии с транспортной функцией, клетки-соседи тесно соприкасаются друг с другом, образуя плотные контакты. Это предотвращает обильное просачивание воды или растворенных веществ с одной стороны эпителиального слоя на другую, минуя клетки.
Образование секрета солевых желез не включает в себя процесс фильтрации, играющий главную роль при образовании мочи в почечных клубочках. Вывод об отсутствии фильтрации можно сделать на основании того, что после введения в кровь мелких фильтрующихся молекул, таких как инсулин или сахароза, в секрете солевых желез они не появляются. Хотя наши знания о работе клеточных механизмов солевой железы скудны, все-таки есть одно надежное свидетельство о существовании в ней активного транспорта NaCl. Na+ - K+ - ATPa3a расположена на базальной мембране эпителиальной клетки, поэтому нанесение в это место уабаина (см. разд. 4.6) блокирует перенос соли. Поскольку указанный ингибитор не проходит сквозь эпителий и способен блокировать На+ - К+-АТРазу только при прямом контакте, активный транспорт Na+, по-видимому, осуществляется в базальной мембране эпителиальной клетки.
Солевые железы у птиц, по-видимому, построены как противоточная система, способствующая
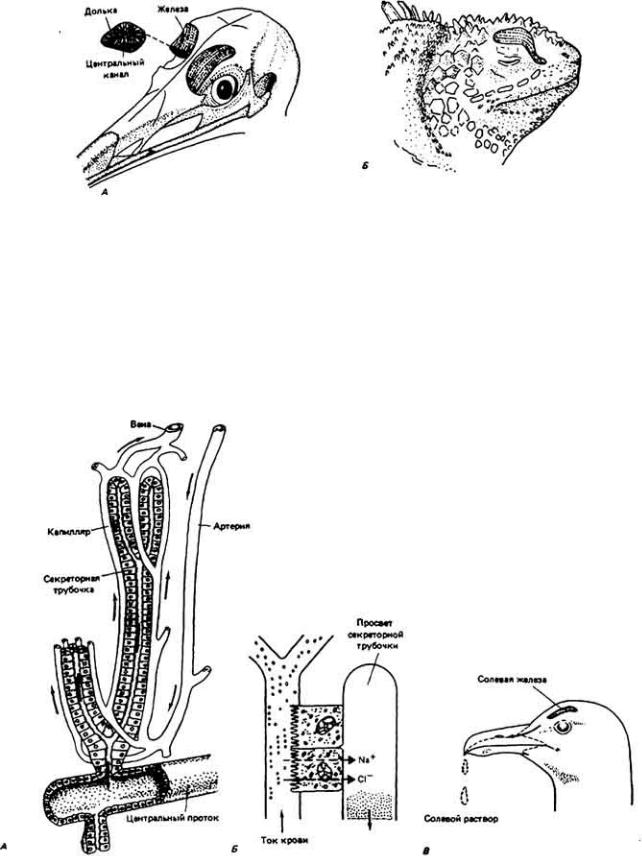
Рис. 12-28.
А . Солевые железы, у птиц расположены над глазницами и соединены через протоки с носовой областью. Железы состоят из множества вытянутых в длину долек. В каждой дольке есть трубки и капилляры (см. рис. 12-29), расположенные по радиусу вокруг центрального канала. Секрет собирается в протоке, впадающем в носовую область. (Schmidt-Nielsen, 1960.) Б. Как установлено, солевые железы в зависимости от вида рептилий могут располагаться возле разных участков тела: глаз, ноздрей, щек, языка. Здесь показана морская игуана, солевая железа которой (обозначено в цвете) находится под кожей. (Dunson, 1969.)
59
Рис. 12-29.
Строение солевой железы птицы. Дольки (не показаны) содержат тысячи разветвленных канальцев, расходящихся по радиусу от центрального протока. А. Каждый отдельный каналец окружен капиллярами, в которых кровь течет в направлении, противоположном току секрета железы в канальце; подобный противоток облегчает перенос соли из крови в каналец. Поскольку величина градиента между концентрациями соли в капилляре и просвете канальца непрерывно уменьшается по мере движения жидкости к центральному протоку
(К. Schmidt-Nielsen. Salt Gland. © 1959. Scientific American Inc. Все авторские права сохранены). Б. Секреторные клетки с обширной щеточной каймой и
митохондриями осуществляют активный перенос Na+ (ионы хлора следуют за
Na+ пассивно). В. Солевой раствор выводится наружу по носовым ходам.
концентрированию солей в выделяемой жидкости. Кровеносные капилляры расположены таким образом, что кровь течет по ним параллельно секреторным трубкам и в направлении, противоположном току выделяемой жидкости (рис. 12-29). Такой характер потоков поддерживает по всей длине трубки минимальный градиент концентрации соли между кровью и секретируемой жидкостью. За счет этого сводится к минимуму градиент концентрации, необходимой для восходящего переноса соли из плазмы крови в секретируемую жидкость.
Секреторная активность солевой железы регулируется прежде всего непосредственно нервной системой (парасимпатической). Вместе с тем существует и нейроэндокринная регуляция (рис. 12-30) данного процесса. При повышении концентрации солей в плазме осморецепторы гипоталамуса реагируют сенсорным возбуждением. Подобная реакция в совокупности с входными нервными сигналами, исходящими из периферических осморецепторов, активирует парасимпатические холинэргические нейроны, которые иннервируют солевые железы. Высвобождаемый из окончаний указанных нейронов ацетилхолин усиливает секрецию соли не только за счет непосредственной стимуляции, но и опосредованно, вызывая расширение сосудов, в результате чего усиливается кровоснабжение секреторной ткани. Гормоны коры надпочечников и пролактин также стимулируют секреторный процесс. Предварительные исследования по эндокринной регуляции солевой железы птиц дают основание полагать, что некоторые гормоны, включая кортикостерон и аргининвазотоцин (АВТ), оказывают регулирующее влияние на секрецию соли. Хотя прямой контроль со стороны нервной системы более важен для быстрой адаптации к осмотическому стрессу, все же для длительного поддержания соответствующей реактивности солевой железы необходим кортикостерон. Например, введение животному с удаленной корой надпочечников (источник кортикостероидных гормонов) гипертонического раствора соли не оказывает стимулирующего эффекта на секреторную функцию солевой железы (рис. 12-31). Но, если затем искусственно ввести в кровь кортикостерон, который не вырабатывается в организме, функция солевой железы восстановится. Было показано, что прямым стимулирующим действием на солевые железы птиц обладает АВТ - пептид, выделяемый ней-рогипофизом в ответ на повышение осмолярности крови и сходный по структуре с АДГ. Железы
60
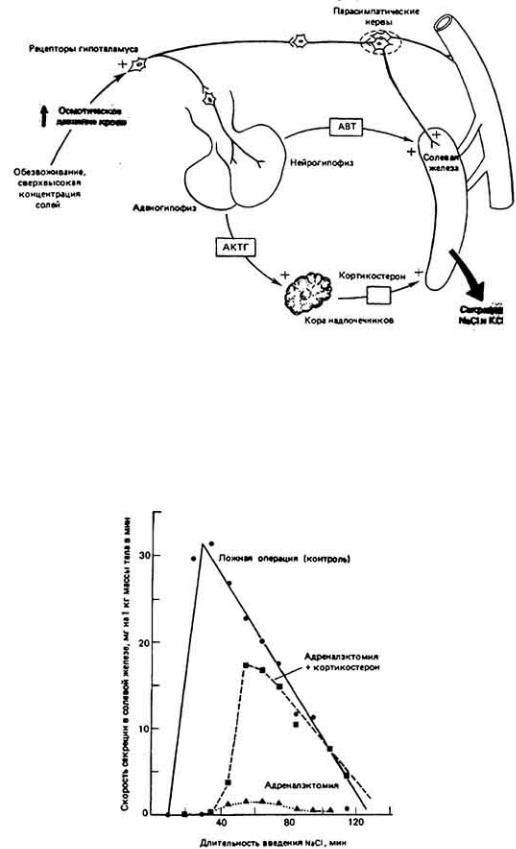
Рис. 12-30.
Упрощенная схема нейрозндокринного регулирования секреции в солевой железе птиц. Осмочувствительные нейроны гипоталамуса в совокупности с входными нервными импульсами из периферических осморецепторов активируют парасимпатические пути иннервации солевой железы и сосудов, снабжающих ее кровью. Таким же образом активируется секреция гипофизом адренокортихотропного гормона (АКТГ), аргининвазотоцина и пролактина (не показано). АКТ Г в свою очередь стимулирует высвобождение из коры надпочечников кортикостерона. (Thomas. Phillips. 1978.)
Рис. 12-31.
Влияние удаления коры надпочечников и картикостероидной заместительной терапии на секреторную функцию солевых желез утки. Через двое суток после операции солевые железы подвергли пробной нагрузке путем введения в кровь 10%-ного раствора NaCl. График, обозначенный темными квадратами, получен при введении NaCl совместно с кортикостероном-стероидным гормоном коры надпочечников. (Thomas, Phillips, 1975.)
отвечают секрецией гипертонической жидкости, в результате чего концентрация солей в крови понижается до значения установочной точки

данной осморегуляторной системы, работающей по принципу обратной связи. Одновременно А ВТ снижает скорость кл у бочковой фильтрации в почках у птиц, которые, как мы ранее отмечали, способны образовывать только слегка гипертоническую мочу.
61
59 :: 60 :: 61 :: Содержание