

Лекція №1
ВВЕДЕННЯ У ВІРУСОЛОГІЮ
План:
1. Зародження та становлення вірусології як науки..
2. Вірусоподібні структури: віріон, віроїд, плазміда, пріон.
3. Значення вірусів у інфекційній патології.
Вірусоло́гія (вірусиі...логія) —наука, яка вивчає властивостівірусівлюдини(медична),тварин(ветеринарна),рослин(сільськогосподарська),бактерій,грибіві процеси, котрі вони спричиняють в організмі чутливих хазяїв, розробляє методи діагностики, лікування та профілактикивірусних інфекцій.
Вивчення властивостей вірусів, особливостей їх взаємодії з клітиною, молекулярно-біологічних механізмів організації таекспресіїгеномувірусів.
Вивчення патогенезувірусних інфекцій, молекулярних механізмів ураження різнихорганіві систем.
Розроблення методів індикації вірусів і методів діагностики вірусних захворювань.
Вивчення закономірностей циркуляції вірусів у різних біоценозах.
Розробка заходів боротьби з вірусними інфекціями та їх профілактики.
Розробка нових біотехнологійзі створення ефективних діагностичних івакцинних препаратів, їх впровадження.
Експериментальне розроблення методів і засобів лікування вірусних інфекцій, їх впровадження.
Слово «вірус» латинського походження і означає «отрута». Вважали, що скажені собаки вводять в організм якусь шкідливу речовину, яку й називали вірусом. Терміном «вірус» користувався Пастер для позначення заразної основи.
Батьком вірусології вважають російського вченого-ботаніка Д.Й. Івановського, який вивчав мозаїчну хворобу тютюну. Розглядаючи у світловому мікроскопі мазки зараженого соку рослин, вчений не виявив жодної бактерії. Тоді в нього виникла ідея пропустити сік рослин через бактеріальний фільтр. Фільтрат не став стерильним, він і далі спричиняв мозаїку в здорових тютюнових рослин.
Івановський, безсумнівно, належить до видатних вчених свого часу: адже йому вдалося зробити не тільки значне відкриття, але і заснувати нову науку - науку про віруси. Розглядаючи під мікроскопом заражені листя рослин, Івановський виявив найдрібніші кристали. Він правильно вирішив, що вони пов'язані з проникненням збудника в рослину. Лише через кілька десятиріччя вчені довели здатність багатьох вірусів формувати кристали всередині заражених клітин при різних захворюваннях не тільки у рослин, а й у тварин.
1898 рік. Німецький мікробіолог Ф. Лефльор разом з П. Фрошем відкрили новий фільтрівний інфекційний агент – вірус ящуру.
1901 рік. Американський військовий лікар В. Рід виявив перший вірус людини – збудник жовтої пропасниці.
1902-1905 роки. Встановлено вірусну етіологію чуми ВРХ, свиней, мясоїдних, віспи, хвороби Ауєскі.
1908 рік. Датські науковці В.Еллерман і О. Банг довели вірусну природу лейкозу птиці.
1911 рік. Американський учений П. Раус відкрив вірус, що викликав у курей розвиток злоякісних пухлин – сарком (його так і називають вірус саркоми Рауса).
У 1917 році канадський мікробіолог Ф. Д'Ерелль відкрив віруси, що паразитують всередині різних бактерій. Д'Ерель назвав їх бактеріофагами, тобто «пожирачами мікробів».
У 1932 році англійський хімік В. Елфорд створює штучні дрібнопористі колоїдні мембрани з точно встановленим розміром отворів в межах від 50 до 300 нанометрів. Пропускаючи через ці мембрани розчини, які містили бактеріофаги і вірус вісповакцини, Елфорд встановлює їх розміри.
1935 рік. Американський вірусолог В. Стенлі виділив вірус тютюнової мозаїки в кристалічному вигляді, за що був удостоєний Нобелівської премії.
1937 рік. Радянські вірусологи Л. Зільбер, Є. Левкович, А. Шубладзе, М. Чумаков, В. Соловйов відкрили вірус кліщового енцефаліту, виявили його переносників – іксодових кліщів, розробили методи діагностики, лікування і профілактики.
1940 рік. Австралійським вченим Вернетом у вірусологічну практику запроваджено використання курячих ембріонів, що значно розширило спектр вірусів, які культивуються в лабораторних умовах.
1941 рік. Американський вірусолог Д. Герст відкрив феномен гемаглютинації, що сприяло дослідженню взаємодії вірусів з чутливими клітинами на моделі вірусу грипу та еритроцитів.
Найбільш швидкий розвиток вірусології почалося після 1948 року, коли Д. Ендерс, відомий американський дослідник-вірусолог, згодом ла уреат Нобелівської премії, розробив метод одношарових тканинних культур.
1949 рік. Американські вірусологи Д. Ендерс, Т. Веллер, Ф. Роббінс встановили здатність вірусу поліомієліту розмножуватися в культурі клітин нирки африканської зеленої мавпи і викликати цитопатичні зміни, за що отримали Нобелівську премію.
В 1952 році, американськими біохіміками Е. Херші та М. Чейз при вивченні бактеріофагів вперше було доведено, що нуклеїнові кислоти відіграють головну роль в репродукції вірусів. На відміну від всіх інших вірусів бактеріофаги не проникають у клітину свого господаря, а лише прикріплюються до його оболонки. Спостерігаючи за допомогою електронного мікроскопа за різними стадіями взаємодії між бактеріофагами і бактеріями, вченим вдалося побачити, як фаг вводить всередину свою нуклеїнову кислоту.
В 1956 році X. Френкель-Конрад в США і одночасно з ним А. Гірер і Г. Шрамм в ФРН зробили важливе відкриття, за яке вони пізніше отримали Нобелівську премію. Зруйнувавши білковий компонент вірусної частинки тютюнової мозаїки карболової кислотою, вони виділили РНК і очистили її. Отримана РНК не містила навіть залишків білка, проте її введення в листя здорових рослин викликало розвиток типової мозаїчної хвороби.
У 1957 році англійський вчений А. Айзекс і його практикантка доктор Д. Лінденман встановили, що якщо внести в культуру тканини інактивований теплом вірус грипу, то клітини починають виробляти білкову речовину інтерферон.
Розшифрували механізм дії інтерферону американський вірусолог С. Барон і радянські вчені, Ф. Єршов і В. Жданов в Інституті вірусології імені Д. Івановського. Вони довели, що процес захисту організму від вірусу відбувається всередині ще не заражених клітин.
1970 рік. Американські вірусологи Г.Тьомін і Д.Балтімор відкрили в складі РНК-вмісних вірусів зворотню транскриптазу – фермент, що переписує генетичну інформацію з РНК на ДНК, за що їх удостоєно Нобелівської премії.
27 жовтня 1979 року в Найробі зібралася Всепланетна комісія, до складу якої увійшли фахівці з різних країн, і проголосили ліквідацію останнього вогнища віспи на нашій планеті.
8 травня 1980 на 33-й сесії ВООЗ була прийнята Декларація глобальної ліквідації віспи. Декларація звернула «увагу на це визначне досягнення до історії охорони здоров'я. Спільні дії науковців усіх країн позбавили людство від хвороби, яка здавна була бідою.
У 1977 році 62 лабораторії світу знищили віруси віспи або передали їх в інші. Ці референс-лабораторії повинні вжити всіх запобіжних заходів, щоб виключити будь-яку можливість винесення вірусу за їх межі. У 1980 року ВООЗ скоротила кількість лабораторій, що зберігають вірус віспи і працюють з ним, до п'яти: в Росії, США, Англії, Нідерландах і Японії.
1982 рік. Американські дослідники Л. Монтанье та С. Прузінер відкрили вірус імунодефіциту людини, що спричиняє СНІД.
1997 рік. Американський вірусолог С. Прузінер встановив етіологію трансмісивних губкоподібних енцефалопатій тварин і людини, за що був удостоєний Нобелівської премії.
2. Віруси – це автонотні генетичні структури,здатні функціонувати і репродукуватися лише в чутливих до них клітинах тварин, рослин, грибів, найпростіших і бактерій. Не будучи організмами, віруси водночас є своєрідною формою життя, зберігають основні атрибути життя, в тому числі здатність до еволюції. Кардинальною відмінністю вірусів від інших організмів, навіть найпримітивніших, є відсутність у них власних білоксинтезувальних систем. Синтез вірусних білків здійснюється на клітинних рибосомах. Тому віруси можуть розмножуватися лише в живих клітинах. Після проникнення віріона в чутливу клітину його оболонки розпадаються, вірусна нуклеїнова кислота звільняється, генетичний апарат клітини блокується, всі інші органели функціонують. Далі на основі записаної на молекулі вірусної ДНК або РНК генетичної інформації клітина зі своїх сировинних матеріалів і своїми системами синтезує вірусні компоненти: нові молекули вірусної нуклеїнової кислоти і вірусних білків.
Окрім класичних вірусів, які представлені віріонами, існують вірусоподібні структури: плазміди, віроїди і пріони.
Плазміди (син.: епісоми, епівіруси) паразитують у цитоплазмі бактерій. На відміну від вірусів, вони не мають білкової оболонки, а складаються лише з молекули кільцевої дволанцюгової ДНК, розміри якої становлять 1 — 3 % розміру геному бактерії. Плазміди можуть вбудуватися в клітинний геном і довготривало перебувати в інтегрованому стані, реплікуючись синхронно з хромосомою. Вони зумовлюють нестабільність генетичних ознак у бактерій. Плазміди виявлено також у дріжджів і плісеневих грибів.
Віроїди – збудники інфекційних хвороб рослин (хризантем, цитрусових, огірків, томатів, картоплі). Як і плазміди, віроїди не мають білкової оболонки, складаються тільки з молекули кільцевої одноланцюгової РНК, яка не кодує жодних білків. Реплікація віроїдів відбувається за участю ферментів рослинної клітини.
Пріони – збудники трансмісивних губкоподібних енцефалопатій тварин і людини, що характеризуються прогресуючим руйнуванням нервових клітин, внаслідок чого головний мозок набуває губчастої структури. Як встановили американські спеціалісти на чолі зі С. Прузінером (1982 - 1993 рр.), ген пріо-на знаходиться в ДНК клітин здорових тварин і людини та продукує нормальний пріонний білок — абсолютно ідентичний патогенному пріону, з тим самим набором амінокислот, тільки з іншою конфігурацією спіралі. Після проникнення в організм інфекційний пріон не розпізнається імунною системою, досягає нейронів, вступає у взаємодію з нормальним пріонним білком, який змінює конфігурацію спіралі й перетворюється на патогенний. Таким чином відбувається ланцюгова реакція, нейрони вакуолізуються і руйнуються, мозкова тканина набуває вигляду губки.
3. Віруси посідають важливе місце в інфекційній патології людини і тварин. За ступенем небезпеки для людей, а також на основі сучасних наукових даних щодо етіології та патогенезу вірусних захворювань,досягнень у профілактиці й лікуванні окремих інфекцій усі віруси, патогенні для людини, поділяють на чотири групи:
I група — збудники особливо небезпечних вірусних інфекцій: віруси Ебола, Марбург, Ласса, Мачупо, натуральної віспи,В-вірус;
II група — збудники висококонтагіозних епідемічних вірусних захворювань: арбовіруси та аренавіруси, які не увійшли до І групи, віруси гепатиту А і В, вірус сказу, вірус імунодефіциту людини;
IIIгрупа— збудники вірусних хвороб, виділених у самостійні нозологічні одиниці: віруси грипу, поліомієліту, групи віспи (за винятком вірусів натуральної віспи і вісповакцини);
IVгрупа — збудники вірусних септицемій, менінгітів, пневмоній, ентеритів: ентеровіруси (за винятком вірусу поліомієліту), рино-, адено-, корона-, реовіруси, віруси парагрипу, паротиту, кору, респі-раторно-синцитіальний, герпесу, вітряної віспи, цитомегалії,червоної висипки, везикулярного стоматиту.
До особливо небезпечних вірусних хвороб тварин (список «А» за даними Міжнародного епізоотичного бюро) належать ящур, нодулярний дерматит ВРХ, чума ВРХ, чума ДРХ, інфекційна катаральна гарячка овець, пропасниця долини Ріфт, африканська чума коней, везикулярний стоматит, везикулярна хвороба свиней, африканська чума свиней, класична чума свиней, віспа овець і кіз, ньюкаслська хвороба, грип (чума) птиці.
У ветеринарній практиці в господарствах промислового типу значно поширені й завдають великих економічних збитків масові респіраторні та шлунково-кишкові захворювання телят, що виникають на фоні стресових факторів і неоднорідного імунологічного статусу. Етіологія респіраторно-кишкових інфекцій телят комплексна. У виникненні їх беруть участь різні віруси (ПГ-3, ІРТ, ВД, РС, адено-, рота-, коро-навіруси та ін.), які спричинюють здебільшого асоційовані інфекції з нашаруванням бактеріальної мікрофлори, що значно ускладнює діагностику і профілактику.
Лекція 2
МОРФОЛОГІЯ, ХІМІЧНИЙ СКЛАД ТА СТІЙКІСТЬ
ВІРУСІВ.
План:
1. Фізична структура вірусів.
2. Нуклеїнові кислоти вірусів
3. Вірусні білки
4. Ліпіди.
5. Вуглеводи.
6. Компоненти клітин у складі вірусів.
7. Стійкість вірусів у зовнішньому середовищі.
ФІЗИЧНА СТРУКТУРА ВІРУСІВ
Віруси існують у двох формах: позаклітинній (віріон) та внутрішньоклітинній (вірусний геном).
Віріон — це метаболічно інертна вірусна частинка, в якій нуклеїнова кислота, що представляє геном (ДНК або РНК), оточена білковою оболонкою — капсидом (від лат. сараа — футляр). Капсид складається а найменших фупкіцонально-еквівалентних структурних одиниць, до складу яких входить одна або кілька молекул білка. Група структурних одиниць утворює морфологічну одиницю — капсомер.
Нуклеїнова кислота разом із капсидом утворює нуклеокапсид. У просто організованих вірусів віріони представлені лише нуклеокапсидами. У складно організованих вірусів нуклеокапсид оточений зовнішньою ліпопротеїновою оболонкою — суперкапсидом, або пеплосом (від грец. реріов — покрив, мантія).
До складу суперкапсиду входять ліпіди і вуглеводи клітини-хазяїна, причому глікопротеїни формують на поверхні віріона морфологічні субодиниці — пепломери, видимі в електронному мікроскопі. Пепломери виглядають як ості (шипи) завдовжки 7 — 10 нм різної форми.
Деякі складно організовані віруси крім суперкапсиду мають ще одну проміжну оболонку — білкову мембрану, утворену М-білком (матриксний, або мембранний, білок), яка оточує нуклеокапсид і формує разом з ним нуклеоїд, або серцевину.
Вірусні капсиди мають надзвичайно впорядковану організацію. Існує два типи будови капсидів віріонів, які забезпечують утворення структур із мінімумом вільної енергії: спіральний та кубічний (ікосаедральний) типи симетрії (рис. 1).
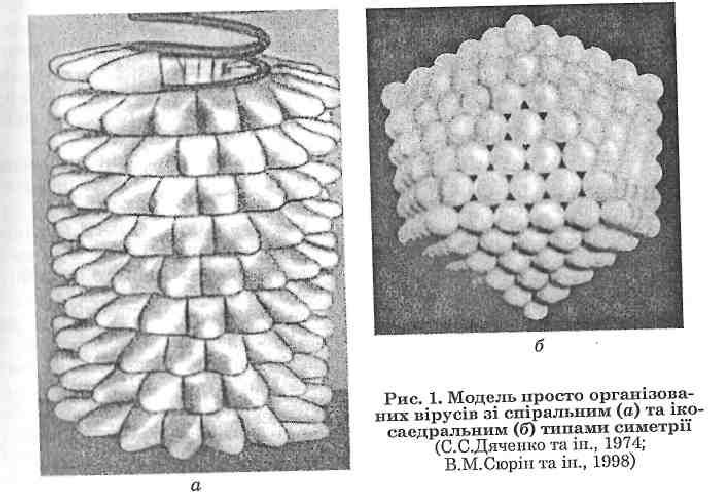
Спіральний тип симетрії характеризується розміщенням капсомерів у вигляді спіралі навколо молекули нуклеїнової кислоти. Віріони зі спіральним нуклеокапсидом мають паличко- або ниткоподібну форму. У разі кубічного (ікосаедрального) типу симетрії капсомери вкладаються у формі геометричної фігури ікосаедра (20-гранника), утворюючи порожнисте ізометричне тіло, в центрі якого знаходиться нуклеїнова кислота.
2. Нуклеїнові кислоти вірусів
На відміну від клітинних форм життя, в складі віріонів є лише один тип нуклеїнової кислоти — ДНК чи РНК. Віруси поділяються на ДНК-вмісні (або ДНК-геномні) та РНК-вмісні (або РНК-геномнї). Вірусна РНК, так само як і ДНК, є носієм генетичної інформації, причому більшість вірусів тварин і людини РНК-геномні. Вірусні нуклеїнові кислоти характеризуються надзвичайною різноманітністю форм. Вірусні ДНК бувають одно- і дволанцюгові, лінійні та кільцеві. У вірусних дволанцюгова ДНУ характеризується тим, що генетична інформація закодована на обох ланцюгах. Структура ДНК в основному унікальна: більшість нуклеотидних послідовностей трапляється лише один раз. Але на кінцях молекул бувають повтори, коли в кінцевому фрагменті ДНК дублюється її початкова ділянка. У цих повторах закладена потенційна здатність до утворення кільцевої форми, яка має велике значення для вірусів.
Вірусні РНК, так само, як і клітинні, дуже різноманітні. Вони бувають одно- і двонитчастими, лінійними, фрагментованими і кільцевими. Для однонитчастої РНК характерна полярність. Віруси, що містять-одноланцюгову РНК, поділяються на дві групи: плюс- нитчасті (віруси з позитивним геномом) і мінус-нитчасті (або віруси з негативним геномом). У плюс-нитчастих вірусів віріонна РНК виконує функцію іРНК, тобто здатна безпосередньо переносити генетичну інформацію на рибосоми. У мінус-нитчастих вірусів віріонна РНК не має властивостей іРНК. У таких вірусів на матриці мінус-нитки РНК синтезується комплементарна їй іРНК за участю вірусного ферменту транскриптази, який обов'язково входить до складу віріонів. Однонитчаста РНК в ортоміксовірусів є фрагментованою ( 7-8 фрагментів). Арена- і буньявіруси також містять однонитчасту фрагментовану РНК (відповідно 2 і 3 фрагменти). Для дволанцюгової РНК характерна фрагментованість. Цей тип нуклеїнової кислоти властивий реовірусам (10-12 фрагментів) і бірнавірусам (2 фрагменти).
Нуклеїнові кислоти зумовлюють інфекційні властивості вірусів.
БІЛКИ
Вірусні білки, як і клітинні, побудовані з амінокислот і являють собою поліпептидні ланцюги, сформовані у вторинні і третинні структури. У зараженій клітині вірусний геном кодує синтез двох групп білків: 1) структурні, які входять до складу віріонів потомства; 2) неструктурні, які забезпечують процес репродукції вірусу на різних етапах, але до складу віріонів не входять.
Структурні білки. Кількість структурних білків вірусів коливається в широких межах і залежить від складності організації віріона. Структурні білки поділяються на дві основні групи: капсидні та суперкапсидні.
Капсидні білки формують капсид, який оточує вірусну нуклеїнову кислоту. Основним принципом будови капсиду є субодиничність. Це означає, що капсидна оболонка складається з білкових субодиниць, які утворені ідентичними поліпептидними ланцюгами. Білкові субодиниці формують капсомери, що розміщені за спіральним або ікосаедральним типами симетрії. Правильно побудовані капсиди віріонів виникають завдяки здатності вірусних білків де самоскладання (впорядкованої агрегації). Самоскладання капсидних білків пояснюється тим, що впорядкована білкова структура — капсид — має мінімум вільної енергії порівняно з невпорядковани-ми білковими молекулами.
Вірусний капсид виконує важливі функції. Основна його функція — захисна, спрямована на захист вірусного геному від несприятливих зовнішніх чинників і насамперед від численних нуклеаз організму. Особливістю капсидних білків є їхня надзвичайно висока стійкість до протеолітичних ферментів. Це досягається завдяки певному укладанню поліпептидних ланцюгів, унаслідок чого пептидні зв'язки стають недоступними для дії ферментів. До складу капсиду просто організованих вірусів входять прикріпні білки, які відіграють важливу роль на початковій стадії взаємодії вірусу з чутливою клітиною. Ці білки розпізнають специфічні рецептори на плазмолемі та забезпечують адсорбцію віріона на клітині. Отже, прикріпні білки здійснюють адресну функцію, що виробилася в процесі еволюції. Завдяки прикріпним білкам вірус розпізнає чутливу клітину, яка здатна забезпечити продукцію повноцінного вірусного потомства. У просто організованих вірусів деякі капсидні білки беруть участь у злитті капсидної оболонки і плазмолеми, що забезпечує проникнення вірусу в клітину. Капсид окремих вірусів (незалежно від складності їхньої організації) містить геномні білки, які ковалентно з'єднані з кінцем вірусної нуклеїнової кислоти. Функції їх нерозривно зв'язані з функціями геному та їхньою регуляцією.
У складі капсиду багатьох складно організованих вірусів містяться ферменти, які беруть участь у репродукції вірусів, зокрема здійснюють транскрипцію та реплікацію вірусного геному
Капсидні білки мають антигенні властивості, тобто індукують синтез специфічних антитіл.
Суперкапсидні білки знаходяться в складі зовнішньої ліпопротеїнової оболонки складно організованих вірусів. Вони пронизують наскрізь подвійний ліпідний шар або не доходять до його внутрішньої поверхні. У більшості вірусів глікопроїни формують на поверхні віріона ості (пепломери), які складаються з кількох молекул ідентичних або різних поліпептидних ланцюгів. Глікопротеїни виконують дві важливі функції, які в кінцевому результаті забезпечують проникнення вірусу в клітину. Одні з глікопротеїнів є прикріпними білками, що взаємодіють із специфічними рецепторами плазмолеми і зумовлюють адсорбцію віріона на клітині, тобто здійснюють адресну функцію. Інші глікопротеїни є білками злиття, що забезпечують проникнення вірусу у клітину шляхом злиття вірусної суперкапсидної оболонки з плазмолемою. Крім того, глікопротеїни є основними антигенами, до яких утворюються вірусонейтралізувальні антитіла. У деяких складно організованих вірусів на внутрішній поверхні суперкапсидної оболонки міститься проміжний структурний шар, сформований матриксним, або мембранним, білком (М-білок). Це гідрофобний білок, який стабілізує структуру віріона та є медіатором складання віріонів потомства, опосередковуючи взаємодію суперкапсидних і капсидних білків.
Неструктурні вірусні білки поділяють на п'ять груп:
І) ферменти синтезу вірусних нуклеїнових кислот ( ДНК і РНК-по-лімерази) - забезпечують транскрипцію та реплікацію вірусного геному;
2) регулятори експресії вірусного геному: мають безпосередній вплив на вірусну нуклеїнову кислоту, запускаючи синтез одних вірусних білків і гальмуючи синтез інших;
3)попередники вірусних білків: відрізняються від інших неструктурних білків нестабільністю в зараженій клітині; з них утворюються структурні вірусні білки в результаті посттрансляційних модифікацій;
4)ферменти модифікацій вірусних білків (протеїнази і протеїнкінази);
5)інгібітори клітинного біосинтезу та індуктори руйнування клітин: руйнують клітинні ДНК та ІРНК; модифікують клітинні ферменти, надаючи їм вірусоспецифічної активності; дестабілізують клітинні мембрани, що зумовлює лізис клітин і звільнення віріонів потомства просто організованих вірусів;
6) нефункціональні пептиди: утворюються в результаті нарізання дуже короткого пептиду, не виконують якоїсь вірусоспецифічної функції.