

Полисомы
|
|
|
|
|
|


В процессе синтеза белка часто образуются так называемые полисомы, или полирибосомы - структуры, состоящие из одной молекулы мРНК и целого ряда связанных с нею рибосом, осуществляющих последовательною интенсивную трансляцию этой мРНК. Полисомы обычно располагаются на поверхности мембран шероховатого эндоплазматического ретикулума(ЭПР), образуя цепочки или спирали. При этом синтезируемый полипептид в результате оказывается внутри полостей ЭПР. Это происходит за счет того, что вначале синтезируется «сигнальная последовательность», богатая гидрофобными аминокислотами. Эта последовательность узнается мембраной ЭПР и связывается с ней. При этом рибосома крепится на мембране с помощью белков рибофоринов, а сигнальная последовательность благодаря своей гидрофобности протискивается сквозь двойной слой фосфолипидов. В дальнейшем рост белковой цепи продолжается, и она постепенно оказывается в полости ЭПР. Этот процесс способствует концентрации белков в замкнутых мембранных полостях, что облегчает их эффективный направленный транспорт в мембранных пузырьках, который осуществляют мембраныкомплекса Гольджис участиеммикротрубочекцитоплазмы.
Вакуоли в дрожжевых клетках

Вакуоли в дрожжевых клетках хорошо видны в световом микроскопе как светлые однородные области внутри клетки. Чаще всего в клетке имеется одна вакуоль, расположенная в центральной части. В процессе почкования вакуоль дробится на более мелкие вакуоли, которые распределяются между клеткой и почкой. Иногда, особенно в удлиненных клетках количество вакуолей достигает 3-4. Основная функция вакуолей в клетках дрожжей сводится к резервному хранению питательных веществ и различных ферментов.
Эндоплазматический ретикулум
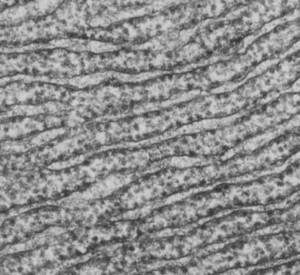
Шероховатый эндоплазматический ретикулум
Эндоплазматический ретикулум (ЭПР) - имеющаяся во всех эукариотических клетках система внутриклеточных мембран, образующих вытянутые мешки, цистерны или узкие каналы. Выделяется два типа ЭПР: шероховатый и гладкий.
Шероховатый, или гранулярный ЭПР покрыт рибосомами, которые на электронно-микроскопических снимках выглядят как многочисленные гранулы. На мембранах рибосомы расположены обычно в виде цепочек и спиралей, образующих полисомы(цепочки рибосом, объединенных одной информационной РНК). Шероховатый ЭПР является важным местом синтеза белков, которые могут накапливаться внутри цистерн и затем включаться в мембранные пузырьки, транспортируемые к месту использования белка (см.комплекс Гольджи).
Гладкий ЭПР не имеет рибосом на поверхности мембран. В цистернах гладкого ЭПР происходят многие метаболические процессы, главным образом связанные с метаболизмом липидов и некоторых внутриклеточных полисахаридов.
Комплекс Гольджи

Структуру, известную как комплекс Гольджи, впервые обнаружил в клетках животных в 1898 г. Камилло Гольджи, итальянский врач и цитолог. Детальное исследование данной структуры сделано позже с помощью электронного микроскопа.
Аппарат Гольджи содержится в цитоплазме почти всех эукариотических клеток, особенно в секреторных клетках животных. У дрожжей комплекс Гольджи выражен несколько хуже, обычно в виде особого отдела эндоплазматического ретикулума. Комплекс Гольджи представляет собой стопку уплощенных мембранных мешочков, так называемых цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи. На одном конце стопки мешочков постоянно образуются новые цистерны путем слияния пузырьков, отпочковывающихся, от гладкого эндоплазматического ретикулума. На другом конце стопки, на внутренней стороне завершается созревание цистерн и они вновь распадаются на пузырьки. Таким образом, цистерны в стопке постепенно перемещаются от наружной стороны к внутренней.
Функцией аппарата Гольджи является транспорт и химическая модификация поступающих в него веществ. Исходным субстратом для ферментов являются белки, поступающие в аппарат Гольджи из эндоплазматического ретикулума. После модификации и концентрирования, ферменты в пузырьках Гольджи переносятся к «месту назначения», например к месту образования новой почки. Наиболее активно этот перенос осуществляется с участием цитоплазматических микротрубочек.
В вакуоляхдрожжевых клеток иногда видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, или волютин. Гранулы состоят в основном из полифосфата:
 .
.
Они служат резервом фосфатных остатков, которые используются для фосфорилирования соединений и синтеза АТФ.