

23. Общие представления о механизме ферментативного катализа. Количественная характеристика ферментативных реакций.
Ферменты — высокоэффективные катализаторы. Они повышают скорость катализируемой реакции в 1012 раз и более.
Кинетика ферментативных реакций - раздел энзимологии, изучающий зависимость скорости химических реакций, катализируемых ферментами, от химической природы реагирующих веществ, а также от факторов окружающей среды. Скорость ферментативной реакции - мера каталитической активности фермента, ее обозначают как активность фермента. Математически скорость ферментативной реакции выражается в изменении концентрации субстрата (уменьшение) или продукта (увеличение) за единицу времени.
К числу главных факторов, влияющих на скорость ферментативных реакций, относятся: концентрация фермента, концентрация субстрата, присутствие ингибиторов или активаторов, рН и температура среды.
Процесс ферментативного катализа условно можно разделить на следующие три стадии:
1) Е + S -— ES (К - k1/k-1) (быстрая) 2) ES-----ЕР (к2)(медленная) 3) ЕР —- Е + Р
Таким образом, в момент равновесия скорости образования и исчезновения комплекса (ES) равны: Е + S — ES-----ЕР — Е + Р
Из
уравнения Михаэлнса следует, что Кт
численно равна конц. субстрата, при
которой скорость реакции равна половине
максимальной; действительно, при условии,
что V= Vmax/2, имеем:
![]()
Более
удобно для определения величин Vmax и Кт
использовать графики линеаризированых
форм уравнения Михаэлиса:
![]()
его
называют уравнением Лайнуивера-Берка.
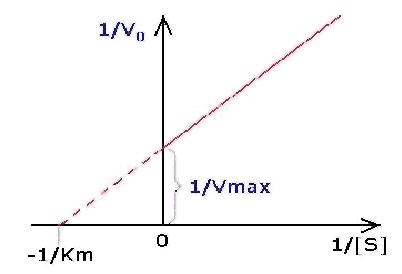
![]()
Для характеристики способности ферментов изменять скорость реакций введено понятие активности. Активность выражают в каталах: 1 кат – количество фермента, которое превращает 1 моль субстрата за 1 с. Другой единицей активности является международная единица (E) – количество фермента, превращающего 1 мкмоль субстрата в 1 мин (1 E = 16,7 нкат).
Ферменты специфически связывают реагенты (свои субстраты) в активном центре. При этом субстраты ориентируются таким образом, что приобретают оптимальное положение для образования переходного состояния. Сближение и необходимая ориентации реагентов значительно повышают вероятность образования продуктивного комплекса. Кроме того, связывание субстрата в активном центре приводит к удалению гидратной оболочки субстрата. В результате удаления молекул воды в активном центре фермента во время катализа создаются совершенно другие условия, чем в растворе. Еще одним важным фактором является стабилизация переходного состояния вследствие взаимодействия между аминокислотными остатками белка и субстратом. Таким образом, переходное состояние в случае ферментативной реакции требует меньшей энергии активации. Кроме того, многие ферменты во время катализа переносят специфические группировки с субстрата или на субстрат. Особенно часто осуществляется перенос протонов. Этот ферментативный кислотно-основной катализ значительно более эффективен, чем обмен протонов с кислотами и основаниями в растворе. Часто химические группировки ковалентно присоединяются к остаткам фермента. Это явление называют ковалентным катализом.