

Эволюционное учение. Горбунов П.С
..pdfМутационный процесс и популяционные волны – факторы-поставщики элементарного эволюционного материала (хотя и действуют по-разному), изоляция оказывается фактором-усилителем генетических различий между группами особей. Механизм действия этих трех основных факторов различен, но общими для них являются ненаправленность, неопределенность и стохастичность действия. Даже совместное действие этих эволюционных факторов не обеспечивает устойчивого протекания направленного процесса эволюции.
Направленность эволюции придает действие естественного отбора.
Словарь-справочник
Борьба за существование – одно из основных понятий в теории эволюции Ч. Дарвина, которое он употреблял для обозначения всей совокупности отношений между особями и различными факторами внешней среды. Эти отношения определяют успех или неудачу данной особи в выживании и оставлении потомства и включают внутривидовую и межвидовую конкуренцию, а также отношения хищник-жертва, взаимодействие организмов с абиотическими факторами внешней среды и др. По Ч. Дарвину борьба за существование – результат, с одной стороны, тенденции организмов к безграничному размножению, с другой – ограниченности природных ресурсов, необходимых для существования особей данного вида. Термин "борьба за существование" Ч. Дарвин предлагал понимать в широком и метафорическом смысле: словом "борьба" он обозначал не столько борьбу, как таковую сколько конкуренцию, а словом "существование" – не только сохранение жизни данной особи, но и успех ее в оставлении потомства. Важнейшей формой борьбы за существование Ч. Дарвин считал внутривидовую конкуренцию, поскольку особи одного вида обладают жизненными сходными потребностями. Борьба за существование приводит к элиминации значительного количества особей вида в каждом поколении и дифференцированному участию в размножении. Результатом борьбы за существование на основе наследственной изменчивости организмов является естественный отбор.
До сих пор идут споры о роли борьбы за существование в эволюции. Ч. Дарвин рассматривал борьбу за существование как предпосылку действия естественного отбора. Объективным единственным мерилом борьбы за существование служит соотношение между числом родившихся особей и числом особей из них, достигших половой зрелости (и соответственно принявших участие в оставлении потомства). Но это не что иное, как естественный отбор – дифференциальное размножение особей в популяциях. Поэтому правильнее считать борьбу за существование не самостоятельным фактором эволюции, а экологической предпосылкой естественного отбора (две другие – наследственная гетерогенность особей в популяциях и давление жизни).
Объективным показателем борьбы за существование является степень расхождения между числом жизней зарождающихся и полностью реализуемых. Простейшие расчеты показывают, что если не возникает препятствий к размножению всех особей, составляющих популяцию, эта популяция будет увеличиваться в геометрической прогрессии, пока не заполнит все пригодное для нее жизненное пространство и пока не будут исчерпаны все доступные ей пищевые ресурсы. Теоретические расчеты подтверждаются наблюдаемыми в природе вспышками массового размножения некоторых животных, растений и микроорганизмов. Такие вспышки всегда вызываются ослаблением борьбы за существование: нарушение стабильности биоценозов (массовое размножение серой зерновой совки в Казахстане при освоении человеком целинных и залежных земель); успешное распространение видов, занесенных извне на новые территории (массовое размножение кроликов в Австралии, одичавших свиней в Новой Зеландии, лошадей в Америке, быстрое распространение завезенной из Америки элодеи в пресных водоемах Европы,
81
нашествие колорадского жука на европейский континент и т.д.).
В 1911 г на о. Сент-Пол у побережья Аляски было завезено 25 северных оленей. Популяция росла по экспоненте до 1938 г., когда на острове площадью 106 квадратных км обитало уже более 2000 оленей. В результате перевыпаса была серьезно подорвана пищевая база (главным образом лишайника) и популяция стала резко сокращаться: в 1950 г. здесь осталось лишь 8 особей.
Формы борьбы за существование
Конституциональная – борьба организмов против неблагоприятных факторов внешней среды (не зависит от плотности популяций).
Внутривидовая прямая (каннибализм в переуплотненных популяциях, активная борьба самцов за самку у полигамных млекопитающих – ластоногие, дикие свиньи, копытные). Внутривидовая косвенная – соревнование, при котором каждая особь стремится обеспечить себя и свое потомство пищей, источниками энергии и жизненным пространством (судьба каждой особи зависит от ее генотипа и экологической обстановки).
Межвидовая прямая – взаимоотношения типа хищник-жертва или паразит-хозяин. Межвидовая косвенная – взаимоотношения определяются принципом конкурентного исключения, когда два вида не могут существовать бесконечно долго в одной и той же местности, если они имеют одинаковые экологические потребности.
Волны жизни, популяционные волны – явление колебания численности популяции. Популяционные волны как общее свойство популяции имеют важное общеэволюционное значение как самостоятельный фактор эволюции. Действие волн жизни как эволюционного фактора предполагает неизбирательную, случайную элиминацию особей, благодаря чему редкий перед колебанием численности генотип (аллель) может сделаться обычным и быть подхвачен в дальнейшем естественным отбором.
В природе встречается множество популяционных волн:
Периодические колебания численности короткоживущих организмов – характерны для большинства насекомых, однолетних растений, большинства грибов и микроорганизмов.
Непериодические колебания численности – зависят от сложного сочетания разных факторов, в первую очередь – от благоприятных для данного вида отношений в пищевых цепях: ослабление пресса хищников для популяций жертв или увеличение кормовых ресурсов для популяций хищников.
Вспышки численности видов в новых районах – территориальные экспансии неизбежно связаны с резким увеличением численности иноземных видов, попадающих
вусловия, благоприятствующие их размножению и расселению главным образом из-за отсутствия естественных врагов: кролики в Австралии, колорадский жук, канадская элодея, американская норка и ондатра в Евразии, африканский подвид медоносной пчелы в Южной Америке, домовая муха около поселений человека и др.
Резкие непериодические колебания численности, связанные с природными катастрофами.
Врезультате популяционных волн происходит случайное колебание концентраций разных генотипов и мутаций – в этом и состоит основное эволюционное значение волн численности. При определенных условиях это случайное и кратковременное колебание численности может превратиться в элементарное эволюционное явление – изменение на протяжении ряда поколений генотипического состава популяции. Влияние популяционных волн особенно заметно в популяциях очень малой численности (не более 500), именно в этих условиях популяционные волны могут как бы подставлять под действие естественного отбора редкие мутации (внося их в увеличенных концентрациях
впопуляционный генофонд) или устранять уже довольно обычные варианты.
Действие популяционных волн статистично и ненаправленно. Популяционные волны служат фактором поставщиком эволюционного материала.
82
Генетическая изоляция – прекращение обмена генами между популяциями близкородственных видов. При генетической изоляции поддерживается обособленность близкородственных видов, предотвращается гибридизация, что способствует созданию большого видового разнообразия в сообществе.
Генетический дрейф (генетико-автоматические процессы) – дрейф генов,
изменение частоты генов в популяции в ряду поколений под действием случайных факторов, приводящее, как правило, к снижению наследственной изменчивости популяций. Наиболее отчетливо проявляется при резком сокращении численности популяции в результате стихийных бедствий (лесной пожар, наводнение и др.), массового распространения вредителей, развития эпизоотии и т.д. Характерная особенность динамики генотипической структуры популяций под действием генетического дрейфа состоит в усилении процесса гомозиготизации, которая нарастает с уменьшением численности популяции. Это нарастание обусловлено тем, что в популяциях ограниченного размера увеличивается частота близкородственных скрещиваний, и в результате заметных случайных колебаний частот отдельных генов происходит закрепление одних аллелей при одновременной утрате других. Некоторые из выщепившихся гомозиготных форм в новых условиях могут оказаться приспособительно ценными. Они будут подхвачены отбором и смогут получить широкое распространение при последующем увеличении численности популяций.
Генетический дрейф – это не целостный фактор эволюции, а совокупность факторов, ибо эффективность генетико-автоматических процессов обусловлена изоляцией, популяционными волнами и мутационным процессом.
Географическая изоляция – географическое разобщение популяций одного вида или близкородственных видов. В условиях географической изоляции происходит аллопатрическое видообразование.
Гетерогенность популяции – генетическое разнообразие особей в популяции. Генетическая гетерогенность природных популяций – главнейшая их особенность. Она поддерживается за счет мутаций, процесса рекомбинации. Генетическая гетерогенность, поддерживаемая мутационным процессом и скрещиванием, позволяет популяции использовать для приспособления не только вновь возникающие наследственные изменения, но и те, которые возникли очень давно и существуют в популяции в скрытом виде. В этом смысле гетерогенность популяций обеспечивает существование мобилизационного резерва наследственной изменчивости.
Границы индивидуальной активности – масштабная характеристика территориально-механической изоляции, зависимая от миграционных способностей видов. Таким образом, степень территориально-механической изоляции определяется не только пространственным расположением соответствующих межпопуляционных барьеров; большое значение имеет репродуктивная активность и подвижность индивидов соответствующего вида.
Изоляция – исключение или затруднение свободного скрещивания между особями одного вида, ведущее к обособлению внутривидовых групп и новых видов. Изоляция – один из элементарных эволюционных факторов. В результате изоляции происходит закрепление начальной стадии дифференциации, консолидации генетического фонда обособившейся группы и становление ее как самостоятельной генетической системы. Все формы изоляции приводят к репродуктивному разобщению, возникновению барьеров между обособившимися видами.
Изоляция как элементарный эволюционный фактор не создает новых генотипов или внутривидовых форм. Значение изоляции в процессе эволюции состоит в том, что она закрепляет и усиливает начальные стадии генотипической дифференцировки, а также в том, что разделенные барьерами части популяции или неизбежно попадают под различное давление отбора. Изоляция ведет к сохранению специфичности генофонда
83
дивергирующих форм. Важнейшая характеристика действия изоляции как фактора эволюции – это ее длительность. В большинстве случаев причина возникновения биологической или пространственной изоляции сохраняется на длительное время. Действие изоляции на эволюционный материал по существу статично и ненаправленно,
ив этом изоляция сходна с другими эволюционными факторами (мутационный процесс
ипопуляционные волны). Изоляция расчленяет исходные популяции на две или более, а группы популяций – на различающиеся формы. Следовательно, действие изоляции – обязательное условие всякого достаточно длительного этапа эволюционного процесса.
Изолирующие механизмы – генетические механизмы (преграды) внешней среды, способствующие уменьшению или блокированию обмена генами между разными популяциями или популяционными системами.
Пространственная (географическая) изоляция – разделение генофонда географическими преградами: предотвращение географическими преградами генного обмена между определенными популяциями. Для пространственной изоляции характерно:
-изоляция какими-либо барьерами между частями видового населения;
-изоляция, определяемая большей возможностью спаривания близко живущих особей, т.е. изоляция расстоянием.
Возникновение пространственной изоляции связано с радиусом репродуктивной активности – степени подвижности особей.
Обыкновенный соловей (Luscinia luscinia), населяющий многие районы европейской территории России, в настоящее время практически сплошь находит подходящие условия для гнездования как в необжитых человеком местах, так и в зарослях по обочинам дорог, в парках и даже скверах больших городов. При этом наблюдается четко выраженная клинальная изменчивость (количественное варьирование признака или комплекса признаков популяций в определенной географической последовательности) пения этих птиц: по числу "коленец", по тембру и другим особенностям существуют постепенные переходы от одной местности к другой (при этом известно, что характер песни наследственно обусловлен). Возникновение такой клинальной изменчивости возможно только потому, что соловьи, несмотря на существование у них частых сезонных перелетов, обладают большим гнездовым консерватизмом: молодые возвращаются практически на то же место, где они вывелись.
Биологическая (репродуктивная) изоляция – изоляция популяций, при которой преграды к скрещиванию порождаются свойствами самих организмов; нескрещиваемость в природных условиях между обитающими вместе организмами.
Этологическая изоляция – психофизиологические, поведенческие различия партнеров в период размножения. Вскрыто большое разнообразие и распространение способов этологической изоляции у животных. Ничтожные на первый взгляд отличия в ритуале ухаживания и обмене зрительными, звуковыми, химическими раздражителями будут препятствовать продолжению ухаживания и вызовут осложнения спаривания.
Морфофизиологическая изоляция – несоответствие в строении и функции половых органов.
Развитие сложных по форме цветов у ряда насекомоопыляемых растений связано с приспособлением цветка к определенному виду насекомых-опылителей. У животных близких видов различия копулятивных органов особенно характерны для некоторых легочных моллюсков, насекомых, а среди млекопитающих – ряда групп грызунов.
Генетическая изоляция – различия наследственного аппарата, приводящие к несовместимости половых клеток, а также гибель зигот после оплодотворения, развитие полностью или частично стерильных гибридов и их пониженная жизнеспособность.
При межвидовом спаривании часто образуются вполне жизнеспособные гибриды, но
уних, как правило, не развиваются нормальные половые клетки. В случае же нормального развития гамет гибриды оказываются малоплодовитыми. В природе есть
84
случаи такой "изоляции посредством гибридизации": на границе обитания двух близких видов постоянно существует зона, населенная вполне жизнеспособными гибридными особями, но их потомство либо ослаблено и не выдерживает конкуренции с более сильными особями родительских видов, либо нежизнеспособно. Такие гибридные зоны известны для некоторых насекомых, серой и черной ворон в Европе.
Сезонная (временная) изоляция – обусловлена различными суточными циклами, изменениями в сроках размножения, сдвигами жизненного цикла изолирующихся полиморфных групп.
Экологическая изоляция – различия сезонных или суточных ритмов в образе жизни и размножении, приспособления к различным участкам биотопа (в вертикальном и горизонтальном направлениях).
У птиц возникают формы, отличающиеся либо сдвигом времени спаривания и гнездования, либо инстинктами гнездостроения, вызывающими локализацию гнезда в разных частях гнездовой стации, характерной для вида, – выше или ниже в кроне дерева, в разных частях кустарничкового яруса, в разных микрочастях луговой или пойменной стации. Так, часть зябликов (Fringilla coelebs) гнездится в Московской области в лесах таежного типа, а другая – в невысоких и редких насаждениях с большим количеством полян (потенциальная возможность перекрестного спаривания особей этих групп ограничена).
Все формы пространственной и биологической изоляции возникают независимо друг от друга и могут сочетаться в любых комбинациях. Возникновению репродуктивной изоляции часто способствует длительная географическая изоляция.
Колебания численности популяции – изменение численности популяции во времени под влиянием абиотических факторов, а также в процессе иммиграции или эмиграции. См. Волны жизни.
Миграции – закономерные перемещения животных между существенно различающимися средами обитания, пространственно отстоящими друг от друга; вызываются изменением условий существования в местах обитания или изменением требований животного к этим условиям на разных стадиях развития (онтогенетические миграции – обеспечивают расселение вида и могут происходить на стадии личинки или во время полового созревания). Миграции животных, вызываемые изменением условий существования в местах обитания животного, могут быть периодическими (сезонные, суточные) и непериодическими.
Периодические миграции – очень разнообразны: вертикальные миграции млекопитающих, птиц и насекомых в горах, беспозвоночных – в толще воды или почвы, проходных рыб – из морей в реки и наоборот, морских черепах и млекопитающих – в океанах, копытных, грызунов, хищных млекопитающих – на суше, многих птиц, летучих мышей, насекомых – в воздухе.
Непериодические миграции – нерегулярные массовые выселения оседлых животных (белок, леммингов, кедровок и др.) под воздействием необычного ухудшения условий (засухи, пожары, наводнения и т.п.), перенаселения; непериодические миграции хаотичны, часто заканчиваются массовой гибелью животных.
Миграции обеспечивают особям возможность выбора в тот или иной момент оптимальных условий существования. Процесс миграции способствует целостности вида и распространению по ареалу наиболее ценных в селективном отношении мутаций. Даже единичные миграции отдельных особей из одной популяции в другую, из зоны обитания одного подвида в зону обитания другого, повторяясь на протяжении сотен и тысяч поколений, создают поток генов. С участием этого механизма регулируется численность особей в популяции и происходит заселение новых мест обитания. Миграции тесным образом связаны с отбором.
Мутационный процесс – постоянно действующий элементарный эволюционный фактор, оказывающий давление на популяции. Мутационный процесс приводит к
85
изменению в популяции частоты одного аллеля по отношению к другому. Хотя по каждому отдельному гену давление мутационного процесса обычно невелико, при наличии большого числа генов в организме оно оказывает заметное действие на генетическую структуру популяции (в сочетании с генетической комбинаторикой). Мутационный процесс и его результат – возникновение мутаций – носят вероятностный и статистический характер. Эволюционное значение мутационного процесса определяется прежде всего тем, что он постоянно поддерживает высокую степень гетерозиготности природных популяций – основу для действия других факторов эволюции и прежде всего естественного отбора. Мутационный процесс – фактор поставщик элементарного эволюционного материала. Мутационный процесс ведет к возникновению части того резерва наследственной изменчивости, который определит в будущем возможность приспособления популяций к тем или иным изменениям условий среды.
Элементарное эволюционное явление – длительное, необратимое и векторизованное изменение популяционного генофонда. Элементарное эволюционное явление – еще не эволюционный процесс. Однако без изменения генотипического состава популяции немыслимо протекание любого эволюционного процесса.
Элементарный эволюционный материал – мутации.
Элементарный эволюционный фактор – см. Естественный отбор. Изоляция.
Волны жизни (популяционные волны). Генетический дрейф (дрейф генов). Мутационный процесс.
Тестовые задания
1.Эволюционные факторы, изменяющие частоты аллелей в популяциях: 1) популяционные волны; 2) наследственность; 3) панмиксия; 4) изоляция; 5) мутационный процесс; 6) конвергенция; 7) борьба за существование; 8) генетико-автоматические процессы
2.Закон Харди-Вайнберга действует при соблюдении следующих условий: 1) панмиксия; 2) размер ареала бесконечно велик; 3) скрещивание особей ограничено; 4) естественный отбор; 5) новые мутации в популяции не возникают; 6) мутационный процесс; 7) миграций особей не происходит.
3.Для осуществления пусковых механизмов эволюционного процесса необходимо наличие фактора: 1) поставляющего в популяцию элементарный эволюционный материал; 2) изменяющего условия существования; 3) расчленяющего популяцию на две или несколько новых; 4) создающего изоляционные барьеры.
4.Элементарным эволюционным материалом является: 1) изменчивость; 2) мутация; 3) наследуемые изменения генетического материала; 4) наследственность; 5) борьба за существование; 6) гетерогенность особей популяции.
5.Эволюционное значение мутационного процесса в природных популяциях: 1) поддерживает высокую степень гетерогенности особей; 2) приводит к ненаследственным изменениям; 3) поддерживает высокую степень гомогенности особей; 4) поставляет элементарный эволюционный материал; 5) приводит к наследственным изменениям.
6.Вспышки массового размножения видов, занесенных извне на новые территории: 1) кролики в Австралии; 2) свиньи в Новой Зеландии; 3) африканизированные пчелы в Южной Америке; 4) одуванчик в Европе; 5) пингвины в Антарктиде.
7.Комбинативная изменчивость связанна с: 1) механизмом распределения хромосом в мейозе; 2) характером протекания эволюционного процесса; 3) случайной встречей гамет при оплодотворении; 4) процессом кроссинговера; 5) изменением условий среды.
8.Факторами-поставщиками элементарного эволюционного материала являются: 1) мутационный процесс; 2) миграции; 3) популяционные волны; 4) изоляция; 5) генетикоавтоматические процессы.
86

9.Колебания численности хищников и их жертв (рис. 49) относятся к элементарному эволюционному фактору – ###.
10.Изоляция как эволюционный фактор: 1) создает новые генотипы; 2) закрепляет начальные стадии генотипической дифференцировки; 3) создает эволюционную независимость изолированных групп;
4)носит направленный характер; 5) приводит к изменению частот аллелей в популяции.
11.Два близкородственных вида американских
Рис. 49. жаб – Bufo americanus и Bufo woodhousii – в тех местах, где они
обитают совместно, часто имеют разные предпочитаемые места размножения. Первый вид откладывает икру в мелкие лужи и ручейки, второй размножается в прудах, болотах и больших дождевых лужах. Это изоляция: 1) морфо-физиологическая; 2) этологическая; 3) биотопическая; 4) пространственная; 5) биологическая.
12.Действие волн жизни как эволюционного фактора предполагает: 1) направленный характер; 2) неизбирательное, случайное уничтожение особей; 3) поставку элементарного эволюционного материала;4) резкое изменение частоты встречаемости аллелей.
13.Популяционные волны случайно и резко изменяют в природных популяциях: 1) морфофизиологическую организацию особей; 2) место обитания; 3) набор мутаций; 4) концентрацию мутаций; 5) условия существования
14.Расселение на северо-восток канареечного
вьюрка (сплошная) и зайца-русака (пунктир) (рис. |
|
|||||
50) |
является |
примером: |
1) |
генетико- |
|
|
автоматических |
процессов; |
2) |
быстрых |
|
||
территориальных экспансий; 3) изоляции; 4) |
|
|||||
популяционных волн; 5) мутационного процесса. |
|
|||||
15. Эволюционное значение популяционных волн |
|
|||||
в природных популяциях: 1) оказывают влияние на |
|
|||||
направление |
и |
интенсивность |
естественного |
|
||
отбора; 2) изменяют концентрацию разных |
Рис. 50. |
|||||
генотипов и |
мутаций; 3) могут |
выводить ряд |
||||
мутаций и генотипов в новую среду обитания для апробации; 4) поддерживают высокую степень гомогенности особей.
16.Собственно генетическая форма изоляции, связанная с возникновением изоляции после оплодотворения, включает 1) гибель зигот после оплодотворения; 2) особое поведение и образ жизни; 3) развитие полностью или частично стерильных гибридов; 4) пониженную жизнеспособность; 5) морфофизиологические особенности
17.Известны следующие формы биологической изоляции 1) физическая; 2) экологоэтологическая; 3) морфофизиологическая; 4) генетическая; 5) географическая
18.Пространственная изоляция может определяться 1) поведением; 2) какими-либо барьерами; 3) морфофизиологической организацией; 4) расстоянием; 5) химизмом среды
19.Соответствие между элементарными эволюционными факторами и примерами
1: мутационный процесс; 2: популяционные волны; 3: изоляция; 4: генетикоавтоматические процессы (дрейф генов); 5: естественный отбор; 6: миграции
А: индустриальный меланизм березовой пяденицы; Б: появление мутации cut («обрезанные крылья») у дрозофилы; В: разорванный ареал голубой сороки в Палеарктике (Европа и Дальний Восток); Г: расширение ареалов на север у вороны, галки, сороки и домового воробья России; Д: популяция эскимосов на севере Гренландии резко отличается по частоте аллелей, определяющих группы крови
87
4.4. Естественный отбор – движущий и направляющий фактор эволюции
Эволюция – направленный процесс, связанный с выработкой новых приспособлений, возникновением одних и вымиранием других видов, возникновением иерархической системы таксонов, осуществлением прогрессивного развития органического мира. Известен лишь один направленный эволюционный фактор – естественный отбор.
Контрольныевопросыизадания
1.Известно, что мутационная изменчивость у организмов, поставляющая материал для естественного отбора, случайна и ненаправленна. Каким образом тогда микроэволюция приобретает направленный характер?
2.Дайте объяснение с эволюционных позиций следующему выражению: «Отбору подвергаются не отдельные гены, а целостные фенотипы. Фенотип выступает не только объектом отбора, но и выполняет роль передатчика наследственной информации в поколениях».
3.У северного подвида узкочерепной полевки, недавно освоившего Полярный Урал, распределение ряда признаков носит асимметричный характер. Например, отмечено увеличение числа эмбрионов у самок, массы печени, сердца и размеров тела. Чем обусловлены такие изменения? О какой форме отбора идет речь в данном случае?
4.Согласно представлениям И.И. Шмальгаузена, любая популяция характеризуется тремя возможными состояниями: неизменность, однонаправленное изменение и разнонаправленное изменение, ведущее к раздроблению. Как эти состояния популяций соответствуют формам естественного отбора? Что можно сказать о генетическом разнообразии популяций во всех трех состояниях?
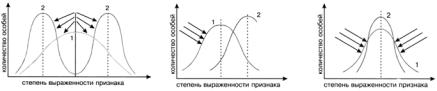
Рис. 51. Схемы действия разных форм естественного отбора
1 – исходная норма реакции; 2 – после действия отбора
5. Американский орнитолог Г. Бампас зимой 1898 г. собрал на улицах Манхэттена 327 домовых воробьев, окоченевших от сильного мороза и метели. Воробьев подвергли биометрическому анализу и
88
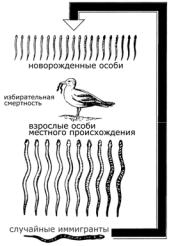
параллельно отогревали в лаборатории. Выжили только те птицы признаки которых приближались к средней норме (длина крыла, длина цевки, масса тела и т.д.). Объясните причину такой элиминации.
6. Поскольку естественный отбор происходит эффективно при достаточно большой численности особей, дальнейшая судьба генофонда малой популяции определяется главным образом действием различного рода случайных факторов. Это было доказано экспериментально С. Райтом. В пробирки с кормом посадили по две самки и два самца мух дрозофил, гетерозиготных по аллелю А (из генотипа Аа). В такой искусственно созданной популяции соотношение нормального и мутантного аллелей было одинаковым (0.5). Спустя 16 поколений оказалось, что оба аллеля остались в 26 популяциях из 96 исходных, в 41 популяции зафиксировался аллель А, в 29 популяциях
– аллель а.
|
7. На островах оз. Эри в Северной |
||||
|
Америке в популяциях ужей по рисунку |
||||
|
поперечных полос встречаются – ужи |
||||
|
белые (А), у них отсутствуют полосы; |
||||
|
ужи темные с наиболее хорошо |
||||
|
выраженными полосами (Г) и ужи с |
||||
|
промежуточной окраской (Б, В) (рис. 52). |
||||
|
Доминирующими |
вокруг |
озера |
||
|
оказываются ужи группы Г, на островах |
||||
|
встречаются группы А, Б и В. |
||||
|
Излюбленными местами обитания ужей |
||||
|
на островах служат известниковые |
||||
|
скалы, обрывы и галечные отмели. |
||||
|
Среди |
новорожденных ужей |
на |
||
|
островах соотношение групп по окраске |
||||
|
было |
следующим: 2А:3Б:12В:4Г, а |
|||
Рис. 52. Естественный отбор |
среди |
взрослых ужей: 4А:4Б:2В:1Г. |
|||
среди ужей озера Эри |
Какие |
формы |
преобладают |
среди |
|
взрослых ужей, а какие среди |
|||||
|
|||||
|
новорожденных? |
|
|
||
Происходит ли избирательная элиминация особей на островах в постэмбриональный период? Почему вероятность выживания полосатых особей на островах примерно 4 раза меньше, чем у особей без полос? Каким могло бы стать соотношение особей разной окраски, если бы прекратилась возможность незначительной миграции материковых ужей на острова, притока аллелей «полосатости» в островные популяции?
89
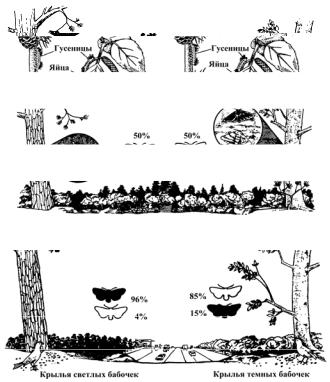
8. В районе Кливленда (Англия) до 1800 г. существовал смешанный лес (сосна, береза, ольха), служивший местом обитания бабочки Oporabia autumnata. Темные и светлые особи этого вида бабочки встречались примерно с одинаковой частотой, и обе формы откладывали яйца как на березе, так и на сосне. Затем смешанный лес был разбит просекой на две части, в результате урагана в южной части леса часть сосен погибло, и вместо них выросли березы; в северной же части леса значительно уменьшилось число растений берез и ольхи.
В XX в. было отмечено, что в сосновом лесу преобладает темная форма, а в березовом – светлая (рис. 53). Продумайте ответы на следующие вопросы. Каким образом в смешанном лесу естественный отбор поддерживал примерно одинаковое соотношение светлых и темных форм бабочек? Как изменилось направление отбора после разделения одного местообитания на два – березовый и сосновый леса? Что может произойти с темными и светлыми формами бабочек в дальнейшем?
Рис. 53. Изменения в популяциях бабочек Oporabia autumnata
90