

Эволюционное учение. Горбунов П.С
..pdfлеса. Клюв тонкий, удобный для добывания семян из еловых шишек. Второй населяет сухие, высокоствольные сосновые леса, но встречается в елово-сосновых лесах, сосняках на болотах. Клюв толстый, массивный, позволяет легко добывать семена из сосновых шишек. Как Вы думаете, что способствовало видообразованию клестов от исходной формы?
Таблица 27.
Изоляция и видообразование
Форма изоляции, путь видообразования
Впопуляциях фиалки часть растений предпочитает кислые почвы, а другая часть – известковые
Впопуляциях пауков самки различным образом реагируют на ритуальные танцы самцов
Различия по форме и размерам цветка, в строении и положении пестиков и тычинок у львиного зева определяют состав опылителей
Существует четко выраженный полиморфизм по размерам
истроению копулятивных органов у жуков-листоедов
Ушироко распространенного на территории Евразии ландыша существуют кавказская, закавказская, европейская, забайальская и дальневосточная популяции
Уястребинки существуют габитуально различные горная, долинная и лесная формы
В пределах обширного ареала бабочка-аполлон образует множество локальных популяций, приуроченных к горным местностям
Умногих растений в популяциях существуют группы особей, различающиеся по форме и размерам куста, типам ветвления, темпу роста и развития. Признаки закреплены генетически.
Убабочек пядениц в популяции существуют меланистические и светлоокрашенные особи
В популяциях насекомых на Гавайских островах есть особи крылатые и бескрылые
4. В плейстоцене обитал древний медведь, от которого произошли травоядный пещерный медведь (он вымер) и современная форма – всеядный бурый медведь. В конце ледникового периода от бурого всеядного медведя произошел белый полярный медведь.
Белые медведи (рис. 62) – самые крупные хищники, масса достигает 1000 кг, длина тела до 3 м. Густая шерсть и подкожный слой толстого жира защищают тело от холода. Белая окраска маскирует хищника, когда он подкарауливает добычу. Жизнь белого медведя связана с морем и плавучими льдами. Основная пища – тюлени, рыба, молодые моржи.
Бурые медведи (рис. 62) обитают в глухих лесах с буреломом; всеядны, имеют очень изменчивую окраску шерсти (от черно-бурой до светло-бурой и
111

светло-серой), имеются отличия и в толщине жирового слоя.
Объясните процесс образования нового вида белого медведя от исходного вида. При объяснении отметьте элементарную эволюционную структуру, элементарный эволюционный материал, поясните значение длительного, направленного изменения генофонда популяций как необходимой предпосылки эволюционного процесса, укажите элементарные эволюционные факторы, под совместным действием которых совершаются элементарные эволюционные явления, приведшие к образованию нового вида медведя.
5. Обитающая в разных районах Евразии обыкновенная лисица образует несколько популяций (рис. 63). Особи северных популяций, распространенные в лесной зоне, самые крупные. Лисы степей и пустынь мельче. Еще мельче лисы, живущие в среднеазиатских пустынях, и самые мелкие лисы обитают в Афганистане, Пакистане, Индии. Замечено, что длина хвоста и ушей также постоянно меняется, но только в обратной зависимости: чем южнее обитают лисы, тем длиннее у них хвост и уши. Объясните возникновение приспособительных изменений у лисы обыкновенной в размерах тела, длине ушей и хвоста в зависимости от среды обитания.
Рис. 63. Подвиды обыкновенной лисицы
Рис. 62. Белый и бурый медведи
(дивергенция видов произошла не более 200 тыс. лет назад)
6. Внимательно познакомьтесь с рисунком 64, проанализируйте события изображенные на нем и ответьте на вопросы. Каковы процессы, в результате которых происходит обособление четырех популяций (a, b, c, d)? Каковы процессы, которые приводят к обособлению нового вида D? Почему не происходит разделение
112
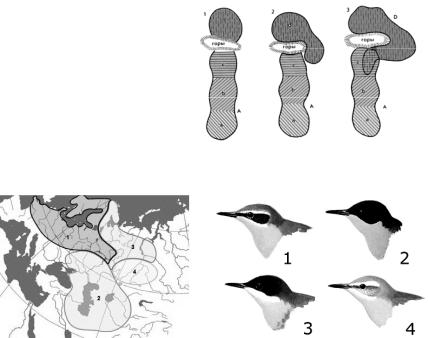
популяций a, b и c на новые виды, хотя и имеются характерные для каждой из них различия? Что происходит в зоне перекрывания ареалов вида A и D?
Вид А – разделен на четыре популяции (a, b, c, d), одна из которых (d) отделена горными хребтами от других популяций;
1 и 2 – начальные стадии видообразования;
3 – обойдя горный хребет с юга, популяция d вновь сблизится с популяцией c, и если они не будут скрещиваться друг с другом, можно считать А и D
самостоятельными видами.
Рис. 64.
7. Выделяют 14 подвидов желтой трясогузки (Motacilla flava), различающихся друг от друга по окраске головы (рис. 65, 66).
Рис. 65. Ареалы желтой трясогузки |
Рис. 66. Подвиды желтой трясогузки |
Каждая из этих форм имеет свой, обычно довольно ограниченный ареал. Как называется вид, включающий в себя несколько подвидов? В каком случае можно этим формам придать статус самостоятельного вида? Какой способ видообразования иллюстрирует данный пример?
8. Вид синица большая представляет сложный комплекс популяций и подвидов, находящихся на разных ступенях изоляции. Ареалы трех подвидов синицы большой – евроазиатского, южноазиатского и восточно-азиатского – образуют кольцо вокруг Центрально-Азиатского нагорья – кольцевой ареал. Подвиды занимают хорошо очерченные ареалы, но в зонах контакта южно-азиатские синицы скрещиваются с двумя другими подвидами. Восточно-азиатские и евроазиатские, обитая совместно в долинах верхнего Амура не скрещиваются. Обособление ареалов синиц и образование их подвидов связано с наступлением ледника.
113
Рассмотрите рисунок (рис. 67). Укажите, какой способ видообразования у синиц. Какой способ видообразования иллюстрирует рисунок 68?
Рис. 67. Ареалы трех подвидов |
Рис. 68. Схема видообразования |
большой синицы |
Эволюционный анализ популяционно-биологических наблюдений
1.На небольших островах в Средиземном море были описаны меланистические расы одного из видов ящериц. Причем, частота темных ящериц была выше на маленьких скалистых островах и меньше на соседних крупных. Темные ящерицы этого вида не встречались на сходных островах (как больших, так и мелких) в Красном море и других более теплых районах. Количество мелких островов с темными ящерицами превышает количество островов со светлыми ящерицами.
Попытайтесь объяснить наблюдаемые явления. Каковы могут быть причины и тип внутривидовой изменчивости ящериц по окраске? Каковы популяции ящериц? Как могло происходить расселение ящериц по островам?
Известно, что темные насекомые быстрее согреваются по утрам и
вхолодные дни. Используйте это для объяснения различий островных популяций ящериц. Укажите направления и формы отбора, действующего в популяциях ящериц. Перечислите другие действующие здесь факторы эволюции.
2.В Австралии кролики, завезенные туда в 1859 году, сильно
размножились и стали вредителями сельскохозяйственных угодий. В 1960 году кроликов с целью истребления заразили вирусом миксомы, вызывающей у них острое заболевание и быструю гибель. В первый же год от миксомы погибло 99% кроликов. Вирус переносится комарами, и летом болезнь распространяется очень быстро. Зимой, когда комаров нет, вирус, не имея промежуточного хозяина, погибает. Каждую весну фермерам приходилось заново интродуцировать вирус. Однако в течение ряда лет гибель кроликов от миксоматоза стала
114
снижаться (в одной из популяций снизилась до 25%). Обнаружилось также наличие у многих кроликов непатогенного вируса миксомы (способного жить и размножаться в организме, не вызывая заболевания и гибели), сохранявшегося в популяции и в зимнее время.
Опишите пути эволюции двух видов – кролика и вируса. В каких направлениях идет в этих популяциях отбор? Какая форма отбора здесь имеется? Каким должен быть общий предполагаемый итог эволюции двух видов?
Что явилось в этом примере элементарным материалом для сопряженной эволюции? Что здесь элементарное эволюционное и адаптационное явления? Каково значение изменений численности популяций кролика и вируса? Все ли кролики (1%), выжившие, не погибшие в первый год борьбы, были устойчивы к вирусу? Можно ли (и как) продолжить борьбу с кроликами, используя вирус миксомы?
3.У серой крысы встречается вариация окраски, называемая "ирландской пегостью" (белое пятно на груди). Анализ географического распространения этого признака показал, что он встречается на всем протяжении евразийского ареала этого вида: от Швеции до Кореи и от Югославии до Камчатки. Частота пегости изменяется от популяции к популяции в широких пределах. Так, на острове Саарема (Эстония) была обнаружена популяция, состоящая исключительно из пегих особей. Часто встречаются популяции, в которых этот признак отсутствует.
Назовите тип изменчивости, который проявляется в данном случае. Как Вы думаете, имеет ли этот признак селективное значение и подвержен ли он отбору? Докажите свою точку зрения. Каким путем возникла своеобразная популяция острова Саарема?
4.В Йоркширском сосновом лесу в 1800 г. произвели вырубку
сосны, и в одной из его частей стала преобладать береза (рис. 53, стр. 83). Части леса разделились широкой просекой. Гаррисон в 1907 г. обнаружил в составе популяций бабочек светлые и темные формы. В сосновой части 96% бабочек темной (меланистической) формы и 4% – светлой. В березовой части 85% – светлой и 15% – темной форм. При изучении крыльев съеденных птицами и летучими мышами бабочек, было обнаружено, что среди съеденных бабочек более 50% светлоокрашенных.
Как изменится соотношение данных окрасок в дальнейшем и какова причина преобладания светлоокрашенных бабочек в березовом лесу? Назовите форму отбора. Можно ли бабочек из березовой и сосновой части леса считать разновидностями? Как поставить опыт по выяснению принадлежности этих бабочек к одной или двум разным популяциям? Можно ли сказать, какая из форм окрасок является доминантной? Является ли
115
отбор фактором, обусловившим появление светлой и темной форм бабочек? Если допустить, что окраски определяются аллелями одного гена, то какова частота этих аллелей в "сосновой" и "березовой" популяциях? Что в данном случае является элементарным адаптационным явлением?
5. Изучение частоты различных вариантов окраски у соболя показало, что темные соболя в 1931-32 годах при очень низкой численности соболя, встречались относительно часто (9.4%). В годы начала восстановления численности (1946-48 гг.) доля темных соболей несколько снизилась (до 5.7%), а в заключительной фазе стабилизации ареала и численности численность темных соболей снизилась до 0.3% (1978 г.).
На зверофермах при попытках получить как можно больше ценных темных соболей в любых скрещиваниях, основная доля потомства была светлой или промежуточной по окраске. Чем объяснить "посветление" природных популяций? Что можно сказать о генотипах светлых и темных соболей? Наблюдается ли здесь элементарное эволюционное явление? Какие действуют факторы эволюции? Какова роль численности? Какие наблюдения и опыты предлагаете Вы поставить, чтобы выяснить причины уменьшения в популяциях частоты темных соболей?
6. В Приангарье с 1974 по 1978 год был изучен фенотип белок из нескольких популяций. Различались два основных варианта окраски – серохвостые и краснохвостые. Численность белок в 1977-78 году была наивысшей, а в 1974-76 гг. наблюдалась депрессия (резкое уменьшение) численности. Было обнаружено, что в одной из популяций (в Жигаловском районе) доля краснохвостых белок в 197778 году по сравнению с 1974-76 гг. возросла с 18 до 42.2%. В двух других популяциях (в Тулунском и Нижне-Удинском районах), доля краснохвостых, наоборот, снизилась до 3.1% и 8.5% соответственно.
Как можно объяснить эти изменения: Назовите форму изменчивости. Какой элементарный эволюционный фактор отвечает в данном случае за изменение частоты признака? Что можно сказать об адаптивной ценности вариантов окраски? Имеется ли в этом примере элементарное эволюционное явление? Как, не проводя скрещиваний, а только наблюдения в природе, и, имея данные за несколько лет, можно уточнить генетическую детерминацию фенотипов белок?
7. На острове Нью-Провиданс (Вест-Индия) улитки различаются по окраске и форме раковины. На западе острова находят улиток с белыми ребристыми раковинами, на востоке преобладают улитки с пигментированными гладкими раковинами. Между этими двумя формами имеется непрерывный ряд переходных форм, промежуточных по окраске и ребристости раковины.
Плате связывает наблюдающиеся различия раковин моллюсков с различиями климата в разных частях острова (на востоке – дождливый, на
116
западе – более сухой), предполагая, что толстая белая раковина лучше предохраняет от перегревания солнечными лучами.
Назовите тип изменчивости, проявляющийся в данном случае. Являются ли эти различия результатом отбора? Если да, то на что действует отбор – на генотип или на фенотип? Какой опыт Вы предполагаете провести для доказательства гипотезы Плате? Как определить экологические и генетические характеристики популяций улитки? Что в данном случае является элементарным адаптационным явлением?
8.На песчаных и илистых грунтах озера Байкал обитают моллюски. В юго-западной части озера они имеют высокую, почти башневидную раковину. Дальше на запад и север раковина становится более низкой и широкой. Вдоль всего северо-восточного побережья раковина моллюсков совсем плоская, с широким пупком. На юго-востоке она вновь преобразуется в высокую. Соседние популяции разделены каменистыми грунтами и частично изолированы. По морфотипу особи соседних популяций почти неразличимы.
Назовите форму изменчивости, проявляющуюся в данном случае. Наследственна ли она? Имеется ли приспособительное значение? Являются ли эти изменения результатом отбора? Как это можно проверить? Имеется ли здесь элементарное эволюционное и адаптационное явление? Как определить экологические и генетические (параметры) характеристики популяций моллюсков?
9.В Северной Америке был описан случай гибридизации в
природе двух видов бабочек-желтушек. В луговых стациях преобладала первая, в посевах люцерны – вторая. Частота гибридов составляла в разных частях ареала от 1 до 12%. Гибриды первого поколения плодовиты и скрещиваются с обеими родительскими формами.
Что можно сказать о происхождении этих двух видов? Как Вы думаете, что можно ожидать в дальнейшем: увеличения или уменьшения частоты гибридов и за счет чего? Почему не происходит полного смешения видов?
10. Якутский заяц-беляк имеет целый ряд морфофизиологических отличий от субтропических и лесостепных популяций того же вида: мелкие размеры, однократное в течение года размножение при большой плодовитости самок, большая амплитуда колебаний численности, способность накапливать жировые запасы.
Объясните, как могла возникнуть каждая из этих черт? Какие из них имеют приспособительное значение для вида (укажите, что выгодно для особи, а что - для популяции в целом)? Попробуйте представить другие возможные пути приспособления популяций того же вида к суровым климатическим условиям. На основе, какой изменчивости формировались указанные отличия зайцев?
117

Результатом микроэволюционных событий, происходящих в популяциях (как элементарных единицах эволюционного процесса), являются формирование элементарных адаптивных явлений и в конце концов видообразование.
Словарь-справочник
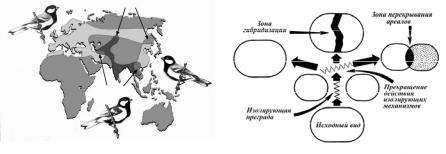
Аллопатрическое видообразование (географическое) – видообразование,
связанное с пространственной изоляцией отдельных популяций данного вида. Популяции, обитающие в различных участках ареала вида, подвергаются действию различных направлений естественного отбора, а географическая изоляция препятствует обмену генетической информацией между обособленными популяциями. Постепенно в таких популяциях происходит генетическая дивергенция, приводящая к микроэволюционным процессам, и особи этих популяций приобретают отчетливые различия с особями исходного вида. Если эти различия приводят к нескрещиваемости с особями других популяций родительского вида, то обособленная форма становится новым самостоятельным видом. «Географическое видообразование – почти единственный способ видообразования у животных и, по всей вероятности, преобладающий способ видообразования у растений».
Возникновение видов ландыша (Convallaria) – исходный родительский вид несколько миллионов лет тому назад был широко распространен в широколиственных лесах Евразии. В четвертичный период произошло сокращение площадей, занятых этими лесами, и единый ареал исходного вида разделился на несколько самостоятельных частей: ландыш сохранился лишь на лесных территориях, избежавших оледенения (на Дальнем Востоке, в Закавказье, Южной Европе). К настоящему времени переживший оледенение на юге Европы ландыш вторично распространился по всей лесной зоне, образовав вид C. majalis. В Закавказье образовался другой вид (C. transcaucasica – более крупный, с широким венчиком), а на Дальнем Востоке – еще один вид (C. keiskii – с красными черешками и восковым налетом на листьях).
Ландыш европейский Ландыш закавказский Ландыш маньчжурский Ландыш японский
Аллопатрия – обитание разных групп организмов (в том числе популяций одного вида)
вразличных географических районах. Действием естественного отбора каждая популяция данного вида приспосабливается к специфическим условиям ее местообитания. Генетически это выражается в присущей каждой популяции характерной частоте встречаемости разных аллелей, а фенотипически – в имеющих приспособительное значение особенностях строения, физиологии, экологии, поведения входящих в ее состав особей. Аллопатрия лежит
воснове аллопатрического видообразования.
Аллопатрические виды – виды с викарирующими ареалами, географически исключающие друг друга. Уровень скрещивания между такими популяциями зависит от пространственного расстояния между ними.
Аллопатрическая гибридизация – гибридизация двух аллопатрических популяций или видов в зоне контакта.
118
Анагенез – способ видообразования во времени, как процесс исторического развития таксономической группы, не сопровождающийся ее распадением на боковые ветви. Способ видообразования, приводящий к исторической смене видов без увеличения видового многообразия. Направление в адаптивных преобразованиях организмов, ведущее к общему усовершенствованию их строения и функционирования и открывающее путь к прогрессивной дальнейшей эволюции всей филогенетической ветви. Ср. Гибридогенез, Кладогенез, Стасигенез.
Изменение раковин моллюсков-палюдин от одного ископаемого вида (Viviparus neymayri) к другому (V. hoernesi) – этот ряд был восстановлен по остаткам раковин, сохранившимся в плиоценовых отложениях Югославии.
Видообразование – процесс возникновения новых видов посредством разветвления предковой филетической линии на несколько новых. Процесс видообразования невозможно понять без выдвинутого Ч. Дарвином принципа дивергенции (расхождения признаков). Дивергенция является неизбежно следствием разнообразия жизненных условий и внутривидовой борьбы за существование. Борьба за существование будет наиболее ожесточенной между представителями того же вида, так как они обитают в той же местности, нуждаются в той же пище и подвергаются тем же опасностям. Особи, уклоняющиеся, т.е. расходящиеся по разным экологическим нишам, смягчают напряженность борьбы. Между специализирующимися крайними формами борьба заметно ослабляется. Отсюда возникает стремление к расхождению признаков, позволяющее занять наиболее разнообразные места в экономике природы. Исходная форма, от которой начинается дивергенция, оказывается в наихудших условиях, так как, не специализируясь в каком-либо направлении, она вынуждена конкурировать с лучше приспособленными крайними формами. Не выдерживая этой конкуренции, промежуточные формы обычно вымирают, и в результате остаются два резко обособленных вида.
Попавшая на небольшой остров в океане первоначально крылатая форма легко уносилась ветром в океан и погибала. Максимальные шансы на выживание имели либо наилучшие летуны, способные противостоять действию ветра, либо лишенные крыльев и вовсе нелетающие насекомые. Этим и определялось расхождение признаков у потомков исходной формы, попавшей на остров. Вместо одного исходного вида могли возникнуть два обособленных: один – с сильно развитыми крыльями, а другой – бескрылый.
Дивергентное видообразование – первоначальным стимулом дивергенции является индивидуальная конкуренция между особями исходного вида, напряженность этой конкуренции снижается по мере того, как вид расчленяется на внутривидовые группы меньшей численности, которые расходятся по разным экологическим нишам и расселяются в пространстве. Такая дифференциация вида уже связана с возникновением межгрупповой конкуренции. В результате осуществляется распределение расходящихся групп (форм) соответственно тем условиям, к которым они в наибольшей степени адаптированы. Прогрессирующее расхождение и специализация потребностей различных групп ведут к дальнейшему ослаблению межгрупповой конкуренции. Каждая форма занимает свое место в экономике природы или полностью прекращается. Исчезает мотив для дальнейшего расхождения признаков, виды стабилизируются, и процесс дивергентного видообразования прекращается.
Таким образом, причинно-следственные связи, определяющие дивергенцию, можно представить следующим образом: Индивидуальная конкуренция ► Расчленение вида на группы ► Расхождение групп по экологическим нишам ► Межгрупповая конкуренция ► Специализация групп, ведущая к образованию новых подвидов и видов ► Прекращение конкуренции и стабилизация видов.
Дивергенция составляет главный способ видообразования.
Пример географической дивергенции – два вида прострела (Anemone pulsatilla): западный (ssp. Germanica) и восточный (ssp. Drandis). Западный подвид имеет тонко рассеченные, разбросанные листья и поникшие цветки. У восточного подвида листья
119

более грубо рассеченные, приподнятые, а цветки стоячие. Этот комплекс изменений связан с обилием дождей на Западе Европы и засушливостью на востоке. Положение листьев способствует стеканию дождевой воды к корневой системе у восточного подвида и, наоборот, разбрызгиванию воды у западного. Поникшее положение цветков предохраняет их от смывания пыльцы дождевой водой. Степень расчлененности листьев тоже имеет адаптивное значение. Она объясняется различной интенсивностью транспирации. Малорасчлененные листья восточного подвида испаряют заметно меньше воды, чем сильно расчлененные листья западного подвида. Различия между этими подвидами прострела носят наследственный характер, хотя пока еще они связаны непрерывным рядом переходных форм.
Следствием многократно повторяющейся дивергенции является адаптивная радиация, когда на основе тех же закономерностей предки одной исходной формы дают пучок расходящихся форм, каждая из которых приспосабливается к определенной нише.
В средней полосе нашей страны произрастает белее двадцати видов лютика. Все они произошли от одного вида. Потомки его заселили различные места обитания – степи, леса, пола – и благодаря изоляции обособились друг от друга сначала в подвиды, потом в виды.
Гибридогенное видообразование – видообразование, связанное с гибридизацией. Межвидовая гибридизация в естественных условиях не такое уж редкое явление, в первую очередь среди растений. Но далеко не всякая гибридизация может породить новый вид. В подавляющем большинстве возникают лишь отдельные гибридные особи, которые либо остаются стерильными, либо не выдерживают конкуренции с особями породивших их видов и быстро исчезают из популяции. Только интрогрессивная гибридизация, происходящая не между единичными особями, а более или менее массовая, может с течением времени и при некоторых условиях привести к зарождению гибридного вида.
Но для этого необходимо, чтобы гибридная форма приобрела способность поддерживать свое существование, сохранять свои отличительные признаки в последующих поколениях и давать плодовитое жизнеспособное потомство либо при однополом размножении, либо при скрещиваниях «внутри себя», т.е. в отсутствие исходных видов. Если это произойдет, мы можем сказать, что в результате гибридогенеза возник новый вид.
Во флоре России интрогессивная гибридизация происходит между даурской и сибирской лиственницами
(Laarix gmelinii и Larix sibirica), между европейской и сибирской елями (Picea abies и Picea obovata).
Культурная слива (Prunus domestica) с 2n=48 возникла путем гибридизации терна (P. Spinosa, 2n=32) с алычой (P. Diivaricata, 2n=16) с последующим удвоением числа хромосом. Некоторые виды пикульников, малины, табаков, брюквы, полыни, ирисов и других растений – также аллополиплоиды гибридогенного происхождения.
Кладогенез – форма эволюции организмов, приводящая посредством адаптивной радиации к образованию из одной группы (таксона) нескольких в пределах прежнего уровня организации. Ср. Анагенез.
Репродуктивная изоляция – защитный механизм вида, который препятствует разрушению его хорошо интегрированной системы коадаптивных генов. Разделение массы разнообразных организмов на виды создает систему, допускающую увеличение генетического разнообразия и накопление выгодных генов и их комбинаций без риска разрушения основного генного комплекса. Репродуктивная изоляция – один из фундаментальных биологических критериев вида.
Симпатрическое видообразование (экологическое) – возникновение новых видов в условиях отсутствия географической изоляции; видообразование, при котором
120