

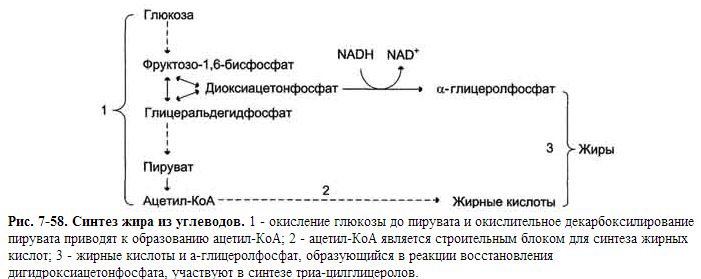
17.Роль "ключевых" ферментов в углеводном обмене.
Гликолиз:
1.гексокиназа:превр глюкозы в глюкозо-6-фосфат,необр реакц,в присут 1АТФ и Мg
2.Фосфофруктокиназа:превр фруктозо-6-фосфат в фруктозо 1,6 бифосфат,необр р,в присут Мg,исп 2АТФ,медлен текущ,опред скорость всей реакц в целом.это аллостерич ферм,ингиб АТФ и Активир АМФ.Акт-ть ФФК ингиб-тся цитратом
3.пируваткиназа:превр фосфоенолпируват в пируват в присутств АДФ,К и Мg
ц.Кребса:
1.цитратсинтаза:ацкоА+оксалоацетат=цитрат
2.изоцитратдегидрогеназа:изолимонн к-та дегидрир и декарбоксилир-тся,НАДзавис.Это аллостерич Е,необх АДФ для ее активац,Mg,Mn
3.альфа-кетоглутаратдегидрогеназа-окислит декарбоксилир альфа-кетоглутарата=сукцинил коА,уч 5 КоЕ:ТПФ,амид липоевой к-ты,НАД,ФАД,НФ-КоА.
Пируватдегидрогеназный комплекс:
1.пируватдегидрогеназа
2.дегидролипоилацетилтрансфераза
3.дегидролипоилдегидрогеназа
+5коЕ: ТПФ,амид липоевой к-ты,НАД,ФАД,НФ-КоА.
Пентозофосфатный путь:генерир НАДН и рибозо-5-фосфат в цитозоле.рибозо-5-фосфат исп в синтезе РНК ДНК
18. Регуляция концентрации глюкозы крови.
Результат регуляции метаболических путей превращения глюкозы - постоянство концентрации глюкозы в крови.
Концентрация глюкозы в артериальной крови в течение суток поддерживается на постоянном уровне 60-100 мг/дл (3,3-5,5 ммоль/л). После приёма углеводной пищи уровень глюкозы возрастает в течение примерно 1 ч до 150 мг/дл
(∼8 ммоль/л, алиментарная гипергликемия), а затем возвращается к нормальному уровню (примерно через 2 ч). На рисунке 7-59 представлен график изменений концентрации глюкозы в крови в течение суток при трёхразовом приёме пищи.
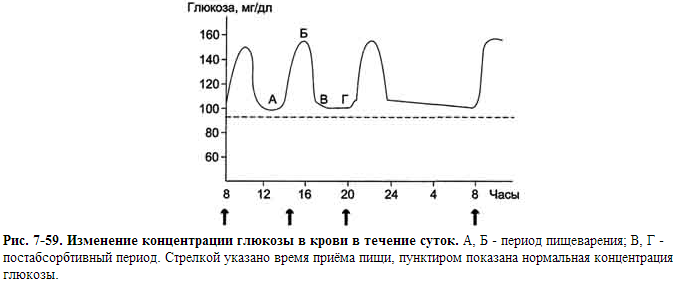
А. Регуляция содержания глюкозы в крови в абсорбтивном и постабсорбтивном периодах
Для предотвращения чрезмерного повышения концентрации глюкозы в крови при пищеварении основное значение имеет потребление глюкозы печенью и мышцами, в меньшей мере - жировой тканью. Следует напомнить, что более половины всей глюкозы (60%), поступающей из кишечника в воротную вену, поглощается печенью. Около 2/3 этого количества откладывается в печени в форме гликогена, остальная часть превращается в жиры и окисляется, обеспечивая синтез АТФ. Ускорение этих процессов инициируется повышением инсулинглюкагонового индекса. Другая часть глюкозы, поступающей из кишечника, попадает в общий кровоток. Примерно 2/3 этого количества поглощается мышцами и жировой тканью. Это обусловлено увеличением проницаемости мембран мышечных и жировых клеток для глюкозы под влиянием высокой концентрации инсулина. Глюкоза в мышцах откладывается в форме гликогена, а в жировых клетках превращается в жиры. Остальная часть глюкозы общего кровотока поглощается другими клетками (инсулинонезависимыми).
При нормальном ритме питания и сбалансированном рационе концентрация глюкозы в крови и снабжение глюкозой всех органов поддерживается главным образом за счёт синтеза и распада гликогена. Лишь к концу ночного сна, т.е. к концу самого большого перерыва между приёмами пищи, может несколько увеличиться роль глюконеогенеза, значение которого будет возрастать, если завтрак не состоится и голодание продолжится
Б. Регуляция содержания глюкозы в крови при предельном голодании
При голодании в течение первых суток исчерпываются запасы гликогена в организме, и в дальнейшем источником глюкозы служит только глюконеогенез (из лактата, глицерина и аминокислот). Глюконеогенез при этом ускоряется, а гликолиз замедляется вследствие низкой концентрации инсулина и высокой концентрации глюкагона (механизм этого явления описан ранее). Но, кроме того, через 1-2 сут существенно проявляется действие и другого механизма регуляции - индукции и репрессии синтеза некоторых ферментов: снижается количество гликолитических ферментов и, наоборот, повышается количество ферментов глюконеогенеза. Изменение синтеза ферментов также связано с влиянием инсулина и глюкагона
Начиная со второго дня голодания достигается максимальная скорость глюконеогенеза из аминокислот и глицерина. Скорость глюконеогенеза из лактата остаётся постоянной. В результате синтезируется около 100 г глюкозы ежесуточно, главным образом в печени.
Следует отметить, что при голодании глюкоза не используется мышечными и жировыми клетками, поскольку в отсутствие инсулина не проникает в них и таким образом сберегается для снабжения мозга и других глюкозозависимых клеток. Поскольку при других условиях мышцы - один из основных потребителей глюкозы, то прекращение потребления глюкозы мышцами при голодании имеет существенное значение для обеспечения глюкозой мозга. При достаточно продолжительном голодании (несколько дней и больше) мозг начинает использовать и другие источники энергии
Вариантом голодания является несбалансированное питание, в частности такое, когда по калорийности рацион содержит мало углеводов - углеводное голодание. В этом случае также активируется глюконеогенез, и для синтеза глюкозы используются аминокислоты и глицерол, образующиеся из пищевых белков и жиров.
В. Регуляция содержания глюкозы в крови в период покоя и во время физической нагрузки
Как в период покоя, так и во время продолжительной физической работы сначала источником глюкозы для мышц служит гликоген, запасённый в самих мышцах, а затем глюкоза крови. Известно, что 100 г гликогена расходуется на бег примерно в течение 15 мин, а запасы гликогена в мышцах после приёма углеводной пищи могут составлять 200-300 г. На рисунке 7-61 представлены значения гликогена печени и глюконеогенеза для обеспечения глюкозой работы мышц разной интенсивности и продолжительности.
Итак, изложенные сведения позволяют сделать вывод о том, что координация скоростей гликолиза, глюконеогенеза, синтеза и распада гликогена с участием гормонов обеспечивает:
-Предотвращение чрезмерного повышения концентрации глюкозы в крови после приёма пищи;
-Запасание гликогена и его использование в промежутках между приёмами пищи;
-Снабжение глюкозой мышц, потребность которых в энергии быстро возрастает при мышечной работе;
-Снабжение глюкозой клеток, которые при голодании в качестве источника энергии используют преимущественно глюкозу (нервные клетки, эритроциты, мозговое вещество почек, семенники).