

Глава 3. Строение глазного яблока
 ного
вещества. Кератоциты обладают большой
степенью
подвижности.
ного
вещества. Кератоциты обладают большой
степенью
подвижности.
Основная функция кератоцитов — синтез межклеточного вещества и коллагеновых фибрилл в период эмбриогенеза, после повреждения роговицы, а также поддержание метаболизма стромы на протяжении всей жизни.
Birk и Trelstad [122] установили, что поверхность фибробластов отвечает за пространственную ориентацию коллагеновых фибрилл. Именно благодаря этому свойству формируются пучки.
В связи с тем, что метаболическая активность кератоцитов в норме снижена, эндоплаз-матическая сеть клеток развита слабо. Лишь после травмы и воспалительных изменений роговицы эндоплазматическая сеть становится хорошо заметной [628].
В строме роговицы встречаются лимфоциты, макрофаги и полиморфноядерные лейкоциты.
Задняя пограничная (десцеметова) пластинка (lamina limitans postrior corneae; Dece-mett). Десцеметова оболочка при световой микроскопии выглядит бесструктурной мембраной, покрывающей заднюю поверхность стромы роговицы (рис. 3.2.3, 3.2.9). В гистогенетическом и структурном смыслах она представляет собой базальную мембрану заднего эпителия роговицы (эндотелия), который ее и продуцирует. Эластичность является одной из наиболее важных ее характеристик. Волокна десцеметовой мембраны образуются на протяжении всей жизни человека. Толщина их при рождении равняется 3 мкм, а в старости — 8—12 мкм [540, 878].
Как и другие базальные мембраны, десцеметова оболочка PAS-положительна и состоит из коротких и тонких фибрилл (10 нм). Фибриллы, в свою очередь, образованы коллаге-
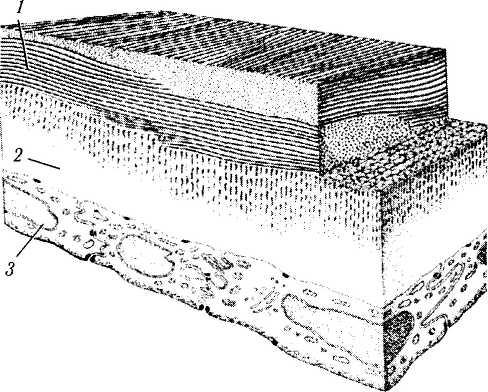
Рис. 3.2.9. Схема микроскопического строения задних слоев роговой оболочки (по Pouliquen, 1969):
1—строма роговой оболочки; 2—десцеметова оболочка; 3— задний эпителий (эндотелий)
ном IV типа и погружены в гликопротеиновое основное вещество [316].
При ультраструктурном исследовании в мембране различают две области [98, 420, 496, 587]. Передняя ее треть имеет толщину 1—4 мкм и задние две трети — 5—15 мкм.
Передний слой десцеметовой оболочки, контактирующий со стромой, имеет многослойный пластинчатый вид, а задний —- гранулированный. Именно передний слой возникает в эмбриональном периоде первым. На тангенциальных срезах этот слой состоит из однородных пластин коллагеновых волокон, образующих равносторонние треугольники. Длина каждой стороны равна ПО нм. Треугольники связаны элек-тронноплотными узлами [1102]. Эти соединения появляются на 5 месяце внутриутробной жизни, когда слой имеют толщину 3,1 мкм (2,2 — 4,5 мкм). Задние 2/з мембраны образуются уже после рождения и состоят из гомогенного фиб-рогранулярного материала.
В мембране, помимо преобладающего коллагена IV типа, обнаружены коллагены III, V, VI и VIII типов [878].
С возрастом в десцеметовой мембране появляются, а затем увеличиваются в количестве коллагеновые волокна и слоистый материал. Этот процесс приводит к появлению на задней поверхности роговицы так называемых бородавок Хассал—Хенле (Hassal—Henle). При этом отмечается нарушение контактов между клетками эндотелия и нарушается барьерная функция последнего.
Несмотря на отсутствие в мембране Десце-мета эластических волокон, она исключительно эластична. При травме нередко десцеметова оболочка скручивается в виде рулона, что обнаруживается при биомикроскопии. Десцеметова мембрана исключительно устойчива в отношении протеолитических ферментов.
Эндотелий (задний эпителий роговой оболочки). Эндотелий роговой оболочки представляет собой один слой плоских гексагональных клеток (плоский однослойный эпителий), расположенных на десцеметовой оболочке (рис. 3.2.3, 3.2.9—3.2.11). Наиболее распространено мнение о том, что они происходят из клеток ней-рального гребня [792, 878, 1105].
Эндотелий роговой оболочки рассматривают как один из наиболее важных структурных компонентов, обеспечивающих прозрачность роговицы [451, 1145]. При этом показано, что обеспечение прозрачности роговицы связано со структурной организацией самой клетки, характера межклеточных контактов и расположением эндотелиальных клеток [128, 260, 261]. Основной функцией эндотелиальных клеток при этом является поддержание постоянного гидростатического давления стромы роговой оболочки. Именно важная роль эндотелия в сохранении прозрачности роговицы явилась причиной многочисленных исследований, направ-
Роговая оболочка и склера
179


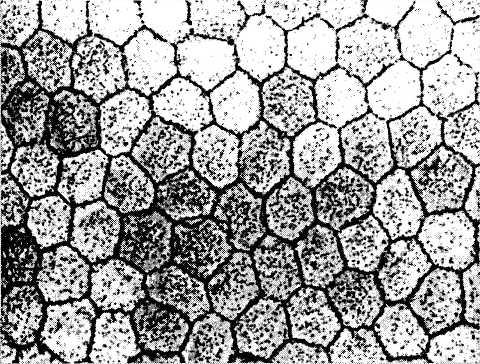
Рис. 3.2.10. Плоскостной препарат эндотелия центральных участков роговой оболочки при исследовании его в фазово-контрастном микроскопе:
отмечается полигональная форма клеток, их примерно одинаковые размеры и наличие плотных контактов между ними
ленных на изучение строения и функции этой структуры глаза. Способствовало этому применение эндотелиальной прижизненной микроскопии.
Последние исследования показали, что у взрослых количество эндотелиальных клеток ограничено и довольно постоянно. Их количество порядка 500 000. С возрастом число клеток уменьшается. Наибольшее уменьшение плотности эндотелиальных клеток определяется в первые годы жизни и полностью коррелирует с увеличением площади роговой оболочки ребенка.
При использовании эндотелиальной микроскопии установлено, что плотность эндотелиальных клеток при рождении колеблется в довольно широких пределах (2627—5316 клеток в мм2) [764]. Плотность клеток падает примерно на 26% на первом году жизни. Дальнейшее падение плотности клеток на 26% отмечается на протяжении последующих 2 лет. Затем скорость уменьшения плотности клеток снижается и число клеток стабилизируется к среднему возрасту [127, 767, 1001]. Кривая, отражающая процесс уменьшения плотности клеток, имеет линейную или логарифмическую форму [262].
В процессе дифференциации уменьшается степень полиморфизма эндотелиального пласта, а также уменьшается количество клеток гексаганальной формы [177, 259, 260, 262, 573, 765, 1113]. Правда, необходимо отметить, что скорость уменьшения плотности и формы клеток колеблется в широких пределах и не дает исследователям возможности сделать окончательное заключение относительно значения этого процесса и факторов, влияющих на этот процесс [1001, 1025].
У молодых людей размер клеток равен 18— 20 мкм (высота — 5—6 мкм), а в более позд-
нем возрасте — 40 мкм [1000]. Появляется би-модальность распределения клеток, как по размерам, так и по содержанию ДНК ядер [36].
Эндотелиальные клетки роговой оболочки присоединяются к десцеметовой оболочке при помощи полудесмосом. Рядом лежащие клетки плотно прилежат друг к другу и соединены десмосомами и запирательными пластинками. Запирательные пластинки распространяются по окружности апикальной поверхности клеток и закрывают межклеточные пространства, обеспечивая барьерные функции эндотелия. Рядом лежащие клетки соединяются также и при помощи «пальцевых вдавлений», представляющих собой цитоплазматические выросты, вдавливающиеся в тело соседней клетки. Несмотря на обилие межклеточных контактов, между клетками существуют щелевидные пространства, шириной 20 нм [163, 487].
Наличие контактов между клетками предопределяет пропускную способность эндотелиального слоя. Они ограничивают пассивный транспорт в строму роговой оболочки. Любое проникновение жидкости в строму через межклеточные щели уравновешивается активным ионным транспортом, происходящим трансцел-люлярно. Процессы регуляции проникновения жидкости в строму могут быть нарушены при уменьшении плотности расположения эндоте-лиоцитов и расширении межклеточных пространств, что нередко наблюдается при патологических состояниях. Рядом исследователей установлены также критические пределы уменьшения количества эндотелиальных клеток. Резкое уменьшение плотности клеток приводит к необратимому нарушению гидратации стромы. Считается, что такой плотностью клеток является величина, равная 400—700 клеток в квадратном миллиметре (при норме 1400— 2500 клеток) [578]. Тем не менее клинические наблюдения показывают, что при ряде патологических состояний даже существенное снижение плотности клеток далеко не всегда сопровождается усилением гидратации стромы роговицы [21].
На апикальной поверхности каждой эндотелиальной клетки располагается от 20 до 30 микроворсинок высотой 0,5—0,6 мкм и шириной 0,1—0,2 мкм. Именно эти образования значительно увеличивают площадь контакта клеточной поверхности с влагой передней камеры глаза. Можно обнаружить и реснички. Они чаще видны по периферии роговицы [889, 918]. Обнаружение ресничек позволило Hogan, Alva-rado, Weddell [496] предположить, что эндотелиальные клетки имеют единое происхождение с клетками трабекулярной сети.
Цитоплазма эндотелиоцитов богата митохондриями, которые обеспечивают энергией активный транспорт, секрецию и высокий уровень синтеза протеинов. Эндотелиоциты содержат митохондрии в значительно большем коли-
180