

Глава 3. Строение глазного яблока
 цитоплазму
клетки условно подразделяют на 3
зоны. Во всех зонах определяется хорошо
развитый
агранулярный эндоплазматический
ретикулум.
цитоплазму
клетки условно подразделяют на 3
зоны. Во всех зонах определяется хорошо
развитый
агранулярный эндоплазматический
ретикулум.
Внешняя треть цитоплазмы эпителиоцитов отличается наличием большого количества митохондрий и складок базальной мембраны. Внутренняя треть цитоплазмы эпителиоцитов насыщена гранулами меланина. Видны также многочисленные свободные и связанные рибосомы. Промежуточная зона цитоплазмы относительно бедна органоидами (рис. 3.6.10). Именно здесь располагается ядро. Комплекс Гольджи выражен нечетко. Его цистерны содержат светлый материал, что свидетельствует о высокой секреторной активности клеток.
Во всех частях цитоплазмы эпителиоцитов располагаются лизосомы обычного строения. Основной их функцией является ферментативное расщепление фагоцитированных фрагментов наружных члеников фоторецепторов [109, 154, 454, 484, 501, 644, 1219, 1971].
Поскольку фагоцитарная активность клеток пигментного эпителия является одной из основных функций [185, 196, 643, 714, 826], их цитоплазма содержит фаголизосомы, образующиеся в результате слияния поглощенных наружных члеников фоторецепторов с первичной лизосомой [524, 1216].
В фаголизосоме первым подвергается лизису белковый компонент фоторецепторных дисков [306, 524, 1216].
Процесс фагоцитоза и лизиса сегментов наружных члеников фотороцепторов происходит довольно быстро. Одна клетка пигментного эпителия кролика в сутки подвергает лизису 2000 дисков в парафовеолярной области сетчатки, 3500 дисков в перифовеолярной области и почти 4000 по периферии сетчатки [484, 1216] (рис. 3.6.11, 3.6.12). Отмечено, что при интенсивном освещении количество фагосом увеличивается. Клетки пигментного эпителия отщепляют наружные членики колбочек таким же образом, как и палочек, но более интенсивно после прекращения освещения [644, 1033]. Процесс разрушения наружных члеников колбочек и палочек фоторецепторов и их утилизации является адаптивным механизмом, способствующим поддержанию структурной и функциональной целостности фоторецепторного аппарата. Тем не менее гибель фоторецепторов возникает также при различных патологических состояниях. Нередко гибель клеток происходит благодаря механизмам апоптоза, находящимся под генетическим контролем [888].
В последнее время проводятся интенсивные исследования роли механизмов апоптоза в развитии большой группы наследуемых дегенеративных заболеваний сетчатой оболочки. Это направление исследований имеет большое практическое значение, поскольку известно более 100 генетически наследуемых синдромов,
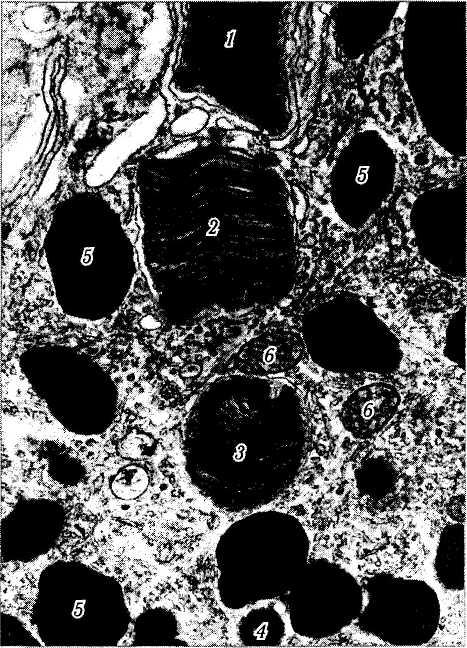
Рис. 3.6.11. Электроннограмма, иллюстрирующая стадии переваривания фрагментов наружных члеников фоторецепторов клетками пигментного эпителия:
/ — наружный членик колбочки; 2 — отделившийся фрагмент наружного членика колбочки и погруженный в цитоплазму клетки пигментного эпителия; 3 — фагосома, содержащая фрагмент наружного членика колбочки; 4 — фагосома на более поздней стадии переваривания фрагмента наружного членика; 5 — мела-носомы; 6 — митохондрии
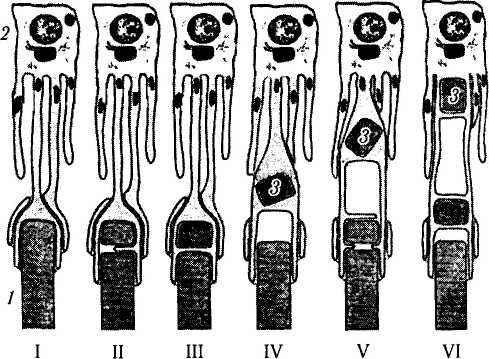
Рис. 3.6.12. Последовательные стадии (I—VI) поглощения и лизиса наружных члеников фоторецепторов пигментными эпителиоцитами сетчатой оболочки. При этом отмечается регенерация наружного членика фоторецептора:
/ — наружный членик фоторецептора; 2 — клетка пигментного эпителия; 3 — фагосома
Сетчатка
235
 сопровождающихся
гибелью нейронов сетчатой оболочки.
Показано, что при некоторых наследуемых
синдромах механизмы апоптоза играют
ведущую роль. При
этом
апоптоз рассматривается
как конечный механизм гибели клеток,
независимо
от характера первичного повреждения.
Основные типы повреждения фоторецепторов
довольно разнообразны и
сводятся
к нарушению их важных функций (синтез
зрительного пигмента, структуры
цитоскелета клеток, последовательности
процессов при восприятии световой
энергии и ее трансформации в нервный
импульс, фагоцитарные функции клеток
пигментного
эпителия и др.) [169, 886—888, 891].
Раскрытие механизмов апоптотической
гибели
нейронов сетчатки и участия в этом
генетического
аппарата рассматривается как наиболее
перспективное направление в лечении
этих заболеваний.
сопровождающихся
гибелью нейронов сетчатой оболочки.
Показано, что при некоторых наследуемых
синдромах механизмы апоптоза играют
ведущую роль. При
этом
апоптоз рассматривается
как конечный механизм гибели клеток,
независимо
от характера первичного повреждения.
Основные типы повреждения фоторецепторов
довольно разнообразны и
сводятся
к нарушению их важных функций (синтез
зрительного пигмента, структуры
цитоскелета клеток, последовательности
процессов при восприятии световой
энергии и ее трансформации в нервный
импульс, фагоцитарные функции клеток
пигментного
эпителия и др.) [169, 886—888, 891].
Раскрытие механизмов апоптотической
гибели
нейронов сетчатки и участия в этом
генетического
аппарата рассматривается как наиболее
перспективное направление в лечении
этих заболеваний.
Нередким структурным включением цитоплазмы клетки пигментного эпителия сетчатки является липофусцин.
Липофусцин содержится во многих тканях организма и его количество нарастает с возрастом. Именно по этой причине этот пигмент был назван «пигментом старения». Возникает он в результате накопления в лизосомах стареющих клеток нелизирующихся агрегатов белка и липидов [1021]. Этот пигмент отличается характерными физико-химическими свойствами, включая естественную желтовато-зеленую флюоресценцию. Накопление липофусцина происходит не только в процессе старения, но и при ряде метаболических заболеваний [1148, 1217]. Причины и механизмы возникновения ли-пофусциноза оставались загадкой более 100 лет. В настоящее время известно, что липофусцин возникает в результате перекисного окисления клеточных компонентов, особенно липидов [1210].
В глазном яблоке, как было указано выше, липофусцин обнаруживается в пигментном эпителии сетчатки [134, 258, 291, 306, 557, 562, 1159, 1176]. Максимальное его накопление происходит в клетках, расположенных в заднем полюсе. К 80 годам липофусциновые гранулы занимают до 19% объема эпителиоцитов [134, 309, 949]. В отличие от других клеток организма, в которых возникает липофусцин в результате аутофагоцитоза внутриклеточных органелл [1021], липофусцин в клетках пигментного эпителия сетчатки возникает в результате фагоцитоза наружных сегментов фоторецепторов [135, 307, 559] с последующим перекисным окислением липидной фракции этих фрагментов. В этом процессе участвует коротковолновой спектр световой энергии [440, 563].
В последнее время указывается на большую роль в формировании липофусцина в эпителиальных клетках сетчатки витамина А и его производных. Об этом свидетельствуют многочисленные экспериментальные биохимические, фи-
зикохимические исследования [291, 292, 558, 559, 561, 1148].
Зерна липофусцина необходимо морфологически отличать от меланосом. Это имеет практическое значение при диагностике пигментных новообразований. Меланиновые гранулы эпителиоцитов имеют круглую или овальную форму. При этом круглые гранулы располагаются в апикальной части клетки, а овальные — в микроворсинках. Липофусциновые гранулы круглые, но менее электронноплотные. Окрашиваются они судановыми красителями и флюоресцируют. Число зерен липофусцина прогрессивно увеличивается с возрастом. Наоборот, количество меланосом с возрастом уменьшается [309, 974, 1159, 1176]. Полагают, что уменьшение количества меланосом связано с деятельностью лизосомного аппарата клеток и возрастным измнением меланина.
Меланин клеток пигментного эпителия поглощает световую энергию достаточно широкого спектра, защищая фоторецепторы и цитоплазму пигментных эпителиоцитов от повреждающего действия света [436]. Меланин обладает свойством свободного радикала и функционирует так же, как полимер, участвующий в обмене электронов. Меланин связывает ряд металлов и лекарственных веществ.
Важно также помнить, что меланиновые гранулы пигментного эпителия сетчатки отличаются от меланосом стромальных меланоцитов уве-ального тракта. Гранулы увеального меланина значительно меньшего размера и имеют овальную форму. Это важно знать патоморфологам, особенно при дифференциальной диагностике внутриглазных пигментных новообразований.
В апикальной части, а также вблизи комплекса Гольджи клеток пигментного эпителия выявляется большое количество пиносом [812]. Размер их меньше (53 нм), чем в эндотели-альных и других клетках (более 100 нм). Эти структуры указывают на наличие интесивных процессов эндоцитоза, характерного для клеток пигментного эпителия.
В цитоплазме эпителиальных клеток можно также обнаружить дискретные темные частицы и пластинчатые тельца. Последние представляют собой фрагменты поглощенных наружных сегментов фоторецепторов [1028, 1219].
3.6.2. Сенсорная часть сетчатки
Сенсорная часть сетчатки представляет собой тонкую прозрачную оболочку, содержащую чувствительные к свету клетки, которые и превращают световую энергию в нервные импульсы. При диафаноскопии глазного яблока сетчатка выглядит пурпурно-красной из-за наличия в фоторецепторах зрительного пигмента (родопсин). Однако этот цвет быстро исчезает при освещении энуклеированного глаза на про-
236