

Глава 3. Строение глазного яблока
 ля
(альдегид витамина А) с опсином
посредством
основания Шиффа. Родопсин относится к
так называемым трансмембранным белкам,
N-конец
которых обращен в междисковое
пространство,
а С-конец обращен в цитоплазму (рис.
3.6.35) [447, 448]. Благодаря этому молекулы
хромофора ориентированы параллельными
рядами вдоль мембран наружных члеников
фоторецепторов,
т. е. располагаются перпендикулярно
падающим на него фотонам, обеспечивая
максимальный сбор световой информации.
Установлено,
что диск наружного сегмента палочки
содержит от 300 до 900 молекул родопсина
[447, 448].
ля
(альдегид витамина А) с опсином
посредством
основания Шиффа. Родопсин относится к
так называемым трансмембранным белкам,
N-конец
которых обращен в междисковое
пространство,
а С-конец обращен в цитоплазму (рис.
3.6.35) [447, 448]. Благодаря этому молекулы
хромофора ориентированы параллельными
рядами вдоль мембран наружных члеников
фоторецепторов,
т. е. располагаются перпендикулярно
падающим на него фотонам, обеспечивая
максимальный сбор световой информации.
Установлено,
что диск наружного сегмента палочки
содержит от 300 до 900 молекул родопсина
[447, 448].
В сетчатке человека выявлено четыре типа зрительных пигментов. Один тип обнаружен в палочках (родопсин) и три в колбочках (иодо-псин). В зависимости от спектральных особенностей поглощения световой энергии колбочко-вые пигменты разделяются на чувствительные к красной (570 нм), зеленой (540 нм) и синей частям спектра (440 нм). 11-цис-ретиналь является хромофором для всех четырех классов зрительных пигментов человека.
Основным механизмом преобразования световой энергии является изменение характера взаимодействия хромофора (11-цис-ретиналь) с белком (опсин). Механизм этого процесса сводится к тому, что при действии световой энергии происходит изомеризация 11-цис-ретиналя с превращением его в полностью транс-рети-наль (рис. 3.6.36). Изменение строения молекулы ретиналя разрушает ее связь с опсином, что приводит к нарушению третичной структуры белка. Этот процесс происходит через ряд звеньев с образованием промежуточных продуктов. Эти промежуточные вещества существуют
11 -цис-ретинил эфир
![]()
11
11-цис-ретинол
ПЭС
П олностью-транс-ре-тинил
эфир
олностью-транс-ре-тинил
эфир
11-цис ретинол__ НАдф
НАДФН + Н+-
![]()
X та О ч
Полностью-транс-ретиналь
1 Опсин
П![]()

 олностью-транс-ретинол
олностью-транс-ретинол
11-цис ретиналь
Родопсин
JI
Н
 СП
СП
Рис. 3.6.36. Химические превращения родопсина в процессе зрительного цикла:
ПЭС — пигментный эпителий сетчатки; НСП — наружный сегмент палочки
исключительно короткое время и их можно анализировать только при низких температурах (рис. 3.6.37). Наиболее важным звеном в этом процессе является переход метародопсина I в метародопсин II. Именно на этом этапе и происходят конформационные изменения белковой части родопсина, что приводит к появлению у последнего ферментативной активности. Эти изменения инициируют дальнейший каскад процессов преобразования, о которых речь пойдет несколько ниже [78, 371, 448].
Родопсин (498 нм)
Свет *- I Пикосекунды
Прелюмиродопсин (батородопсин) (543 нм)
I Наносекунды
Люмиродопсин (497 нм)
I Микросекунды
Метародопсин I (478 нм)
I Миллисекунды
Метародопсин II (380 нм)
I Секунды
Метародопсин III (465 нм)
I Минуты
Опсин (280 нм)
+ Транс-ретиналь (380 нм)
Рис. 3.6.37. Схема превращений родопсина под действием световой энергии (в скобках указаны спектральные изменения продуктов реакции)
После разрушения связи хромофора с опсином наступает обратный процесс, т. е. регенерация родопсина. Происходит это следующим образом (рис. 3.6.36). При обесцвечивании зрительного пигмента полностью-транс-ре-тиналь высвобождается из зрительного пигмента и преобразуется в полностью-транс-ретинол. Полностью-транс-ретинол из наружных сегментов фоторецепторов поступает в пигментный эпителий сетчатки, где он эстерифицируется, превращаясь в эфир полностью-транс-ретинил эфир. Последний превращается в 11-цис-ретинол благодаря деятельности фермента — рети-ноид изомеразы. Образовавшийся в результате реакции 11-цис-ретонол возвращается в фоторецепторы, где, окисляясь, превращается в 11-цис-ретиналь. 11-цис-ретиналь соединяется с опсином, образуя родопсин. Вновь образованный родопсин может опять абсорбировать
Сетчатка
253

Темновые
условия
Наружный сегмент Внутренний
сегмент
3Na*
-No*
Свет
Из приведенной цепи реакций видно, что составленные части родопсина повторно используются в зрительном цикле. Тем не менее процесс регенерации хроматофора предполагает обязательное постоянное пополнение клеток пигментного эпителия витамином А, из которого образуется эфир 11-цис-ретинила.
В организм человека витамин А поступает с пищей и хранится в печени. Поступая в кровь, он связывается с ретинол-связывающим белком и затем с преальбумином. Этот белковый комплекс, благодаря наличию фенестр в эндотели-альной выстилке капиллярных сосудов хориои-деи, легко проникает через мембрану Бруха и достигает клеток пигментного эпителия сетчатки. Затем витамин А отделяется от белковой части комплекса и поступает в цитоплазму пигментных клеток для дальнейших преобразований в 11-цис-ретиналь.
Для восстановления родопсина необходимо пополнение и его белковой части, т. е. опсина. Пополнение фоторецепторов опсином происходит благодаря постоянно протекающему процессу регенерации наружных члеников палочек и колбочек. Вновь образованные мембранные пакеты, содержащие в своем составе и опсин, постепенно передвигаются к апикальной поверхности фоторецептора, где опсин связывается с 11-цис-ретиналем, образуя «новый» родопсин.
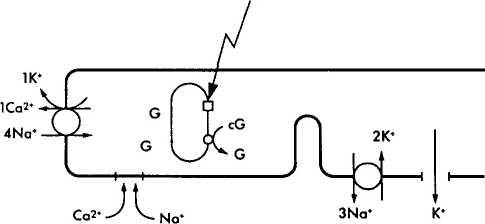
Кратко описав характер химических преобразований родопсина в процессе зрительного цикла, необходимо ответить на вопрос — каким образом описанные физико-химические процессы приводят к инициализации нервного импульса? Чтобы понять этот процесс необходимо обратиться к рис. 3.6.38. На рисунке видно, что в темновых условиях фоторецепторы деполяризованы. Это связано с тем, что натриевые каналы плазматических мембран сегментов палочек и колбочек в темноте открыты и из внеклеточного пространства в цитоплазму фоторецептора поступает большое количество ионов натрия. При этом диффузия натрия из наружного сегмента фоторецепторов во внутренний сегмент в темновых условиях обеспечивает формирование «темнового тока» [1206].
Натриевые каналы остаются открытыми благодаря высокой концентрации циклического гуанозин монофосфата (cGMP). Равновесие между ионами натрия и калия поддерживается благодаря деятельности АТФ-зависимого натрий/калиевого насоса.
Воздействие на зрительный пигмент световой энергии приводит к закрытию ионных каналов и снижению проводимости Na+ через мембрану наружного сегмента (рис. 3.6.38). При этом изменяется трансмембранный потенциал фоторецептора и возникает гиперполяризация.
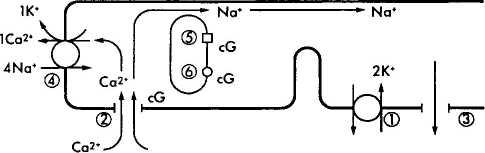
Рис. 3.6.38. Схематическое изображение механизма
формирования нервного импульса в фоторецепторной
клетке:
В темноте ионы натрия (Na+), как и ионы кальция (Са+), перемещаются из наружного сегмента фоторецептора во внутренний благодаря деятельности Na+/K насоса (/), а поступают в наружный сегмент через катионные каналы (2). При этом формируется «темновой ток» ионов натрия. Катионные каналы открыты тогда, когда сСМР (cG) находится в связанном состоянии. Поток ионов натрия в направлении внутреннего сегмента происходит по мере выхода из клетки ионов калия (3). Вследствие поглощения фотона родопсином (5) активизируется фосфодиэстераза (6), что приводит к повышению концентрации cGNP и закрытию катионных каналов. Следствием этого является уменьшение проницаемости мембраны для ионов натрия и усиление ее поляризации. Посредником в этом процессе являются ионы кальция, поскольку они эффективно блокируют натриевые каналы и вызывают наблюдаемую гиперполяризацию. Выведение ионов кальция обеспечивается деятельностью ионообменника (4) и при закрытых каналах
Таким образом, фоторецепторы отвечают на освещение не потенциалами действия, а гиперполяризацией, величина которой пропорциональна интенсивности освещения.
В деполяризованном (темновом) состоянии фоторецепторы высвобождают нейромедиаторы в синаптическую щель, которые взаимодействуют с постсинаптическими терминалами биполярных и горизонтальных клеток.
Увеличение степени освещенности вызывает градуированную гиперполяризацию, которая вызывает уменьшение выделения нейромедиа-тора.
Необходимо отметить, что фоторецепторы, как и горизонтальные и биполярные клетки, не генерируют потенциалы действия, и таким образом отвечают на световую энергию уменьшением выделения медиатора [1206, 1055]. Только нейроны третьего порядка (ганглиозные клетки) генерируют потенциалы действия.
Как указано выше, индуцирует гиперполяризацию фоторецептора перекрытие ионных каналов. Изучению механизмов этого процесса
254