

Глава 3. Строение глазного яблока

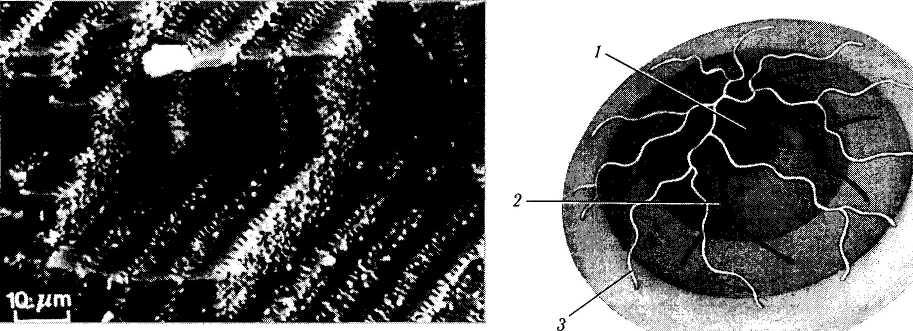
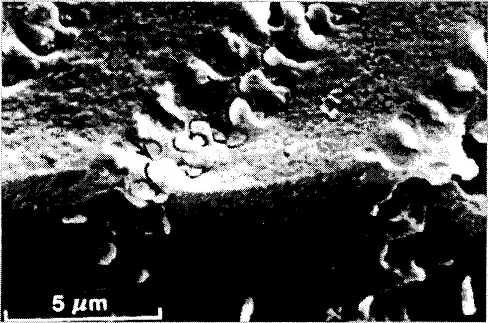
Рис. 3.4.12. Характер расположения хрусталиковых
волокон. Сканирующая электронная микроскопия
(по Kuszak, 1989):
а—плотно упакованные хрусталиковые волокна; б — «пальцевые вдавления»
По мере погружения к центру хрусталика постепенно нарушается их однообразие по размеру и форме. В области экватора у взрослых ширина хрусталикового волокна колеблется от 10 до 12 мкм, а толщина — от 1,5 до 2,0 мкм. В задних частях хрусталика волокна более тонкие, что объясняется асимметричной формой хрусталика и большей толщиной передней коры [621, 624]. Длина хрусталиковых волокон в зависимости от глубины расположения колеблется от 7 до 12 мм [183]. И это при том, что первоначальная высота эпителиальной клетки равняется всего 10 мкм.
Концы хрусталиковых волокон встречаются в определенном месте и формируют швы.
Швы хрусталика (рис. 3.4.13). В феталь-ном ядре имеется передний вертикально расположенный Y-образный и задний инвертированный Y-образный швы. После рождения по мере роста хрусталика и увеличения количества слоев хрусталиковых волокон, формирующих свои швы, происходит пространственное объединение швов с образованием звездоподоб-ной структуры, обнаруживающейся у взрослых.
Рис. 3.4.13. Формирование швов в месте стыка волокон, происходящее в различные периоды жизни:
/ — Y-образный шов, формирующийся в эмбриональном периоде; 2 — более развитая система швов, возникающая в детском периоде; 3 — наиболее развитая система швов, обнаруживаемая у взрослых
Основное значение швов заключается в том, что благодаря такой сложной системе контакта между клетками сохраняется форма хрусталика практически на протяжении всей жизни.
Особенности мембран хрусталиковых волокон. Контакты типа «пуговица — петля» (рис. 3.4.12). Мембраны соседствующих хрусталиковых волокон соединены при помощи разнообразных специализированных образований, изменяющих свое строение по мере смещения волокна с поверхности в глубь хрусталика. В поверхностных 8—10 слоях передних отделов коры волокна соединяются при помощи образований типа «пуговица — петля» («шар и гнездо» американских авторов), распределенных равномерно по всей длине волокна. Подобного типа контакты существуют только между клетками одного слоя, т. е. клетками одного поколения, и отсутствуют между клетками разных поколений. Это обеспечивает возможность передвижения волокон относительно друг друга в процессе их роста.
Между более глубоко расположенными волокнами контакт типа «пуговица — петля» обнаруживается несколько реже. Распределены они в волокнах неравномерно и случайным образом. Появляются они и между клетками различных поколений.
В самых глубоких слоях коры и ядра, кроме указанных контактов («пуговица — петля»), появляются сложные интердигитации в виде гребней, впадин и борозд [629, 798, 1170]. Обнаружены также и десмосомы, но только между дифференцирующимися, а не зрелыми хруста-ликовыми волокнами.
Предполагают, что контакты между хрус-таликовыми волокнами необходимы для поддержания жесткости структуры на протяжении всей жизни, способствующей сохранению про-
Хрусталик и ресничный поясок (зонулярный аппарат)
213
 зрачности
хрусталика. Еще один тип межклеточных
контактов обнаружен в хрусталике
человека.
Это щелевой контакт [825]. Щелевые контакты
выполняют две роли. Во-первых, поскольку
они соединяют хрусталиковые волокна
на большом протяжении, сохраняется
архитектоника
ткани, тем самым обеспечивается
прозрачность
хрусталика [625]. Во-вторых, именно
благодаря наличию этих контактов
происходит распространение питательных
веществ между
хрусталиковыми волокнами. Это особо
важно
для нормального функционирования
структур
на фоне пониженной метаболической
активности
клеток (недостаточное количество
органоидов).
зрачности
хрусталика. Еще один тип межклеточных
контактов обнаружен в хрусталике
человека.
Это щелевой контакт [825]. Щелевые контакты
выполняют две роли. Во-первых, поскольку
они соединяют хрусталиковые волокна
на большом протяжении, сохраняется
архитектоника
ткани, тем самым обеспечивается
прозрачность
хрусталика [625]. Во-вторых, именно
благодаря наличию этих контактов
происходит распространение питательных
веществ между
хрусталиковыми волокнами. Это особо
важно
для нормального функционирования
структур
на фоне пониженной метаболической
активности
клеток (недостаточное количество
органоидов).
Выявлено два типа щелевых контактов — кристаллические (обладающих высоким омическим сопротивлением) и некристаллические (с низким омическим сопротивлением). В некоторых тканях (печень) указанные типы щелевид-ных контактов могут преобразовываться один в другой при изменении ионного состава окружающей среды. В волокне хрусталика они неспособны к подобному преобразованию [392] Первый тип щелевых контактов найден в местах прилегания волокон к эпителиальным клеткам, а второй — только между волокнами [106, 627].
Низкоомные щелевые контакты содержат внутримембранные частицы, не позволяющие соседним мембранам сближаться более чем на 2 нм. Благодаря этому в глубоких слоях хрусталика ионы и молекулы небольшого размера достаточно легко распространяются между хрусталиковыми волокнами, и их концентрация довольно быстро выравнивается. Имеются и видовые различия в количестве щелевых контактов. Так, в хрусталике человека они занимают поверхность волокна по площади 5%, у лягушки— 15%, у крысы — 30%, а у цыпленка — 60% [625, 626, 665]. Щелевых контактов нет в области швов.
Необходимо кратко остановиться на факторах, обеспечивающих прозрачность и высокую рефракционную способность хрусталика. Высокая рефракционная способность хрусталика достигается высокой концентрацией белковых филаментов, а прозрачность — их строгой пространственной организацией, однородностью структуры волокон в пределах каждого поколения и небольшим объемом межклеточного пространства (менее 1% объема хрусталика). Способствует прозрачности и небольшое количество внутрицитоплазматических органоидов, а также отсутствие в хрусталиковых волокнах ядер. Все перечисленные факторы сводят к минимуму рассеивание света между волокнами.
Есть другие факторы, влияющие на рефракционную способность. Одним из них является увеличение концентрации белка по мере приближения к ядру хрусталика. Именно благодаря увеличению концентрации белка отсутствует хроматическая аберрация.
Не меньшее значение в структурной целостности и прозрачности хрусталика имеет и регуляция ионного содержания и степени гидратации волокон хрусталика. При рождении хрусталик прозрачен. По мере роста хрусталика появляется желтизна ядра. Возникновение желтизны, вероятно, связанно с влиянием на него ультрафиолетового света (длина волны 315—400 нм). При этом в коре появляются флюоресцирующие пигменты. Предполагают, что эти пигменты экранируют сетчатку от разрушительного действия коротковолновой световой радиации [1011]. Пигменты накапливаются в ядре с возрастом, а у некоторых людей участвуют в образовании пигментной катаракты. В ядре хрусталика в старческом возрасте и особенно при ядерной катаракте увеличивается количество нерастворимых белков, которые представляют собой кристаллины, молекулы которых «сшиты».
Метаболическая активность в центральных участках хрусталика незначительна. Практически отсутствует обмен белков [446]. Именно поэтому они относятся к долгоживущим белкам и легко подвергаются повреждению окислителями, приводящими к изменению конформации белковой молекулы из-за образования сульф-гидрильных групп между молекулами белка. Развитие катаракты характеризуется увеличением зон рассеивания света. Это может быть вызвано нарушением регулярности расположения хрусталиковых волокон, изменением структуры мембран и нарастанием рассеивания света, в связи с изменением вторичной и третичной структуры белковых молекул. Отек хрусталиковых волокон и их разрушение приводит к нарушению водно-солевого обмена.