

1.1.3. Структура бактериальной клетки и методы ее исследования
Бактерии являются прокариотами (см. рис. 1.2) и существенно отличаются от клеток растений и животных (эукариотов). Они относятся к одноклеточным организмам и состоят из клеточной стенки, цитоплаз- матической мембраны, цитоплазмы, нуклеоида (обязательных компонентов бактериальной клетки). Некоторые бактерии могут иметь жгутики, капсулы, споры (необязательные компоненты бактериальной клетки).
(запасные
вещества)
мембранные
структуры структуры
Рис.
1.2. Комбинированное схематическое
изображение прокариотиче- ской
(бактериальной) клетки со жгутиками.
1
- гранулы полиоксимасляной кислоты; 2
- жировые капельки; 3 - включения
серы; 4 - трубчатые тилакоиды; 5 -
пластинчатые тилакоиды; 6 - пузырьки;
7 - хроматофоры; 8 - ядро (нуклеоид); 9 -
рибосомы; 10 - цитоплазма; 11 - базальное
тельце; 12 - жгутики; 13 - капсула; 14 -
клеточная стенка; 15 - цитоплазматическая
мембрана; 16 - мезосома; 17 - газовые
вакуоли; 18 - ламеллярные структуры; 19
- гранулы полисахарида; 20 - гранулы
полифосфата
Включения Нефотоситезирцющие Основные
Клеточная стенка
Клеточная стенка представляет собой внешнюю структуру бактерий толщиной 30-35 нм, главным компонентом которой является пеп- тидогликан (муреин). Пептидогликан является структурным полимером, состоящим из чередующихся субъединиц N-ацетилглюкозамина и N- ацетилмурамовой кислоты, соединенных гликозидными связями (рис. 1.3).
Рис.
1.3. Схематическое изображение однослойной
структуры пептидо-
гликана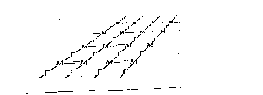
Параллельно расположенные полисахаридные (гликановые) цепи скреплены между собой поперечными пептидными мостиками (рис. 1.4).
Рис.
1.4. Детальное строение структуры
пептидогликана Светлые
и черные короткие стрелки указывают
связи, расщепляемые соответственно
лизоцимом (мурамидазой) и специфической
муроэндопептидазой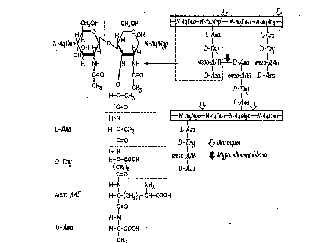
Полисахаридный каркас легко разрушается лизоцимом - антибиотиком животного происхождения. Пептидные связи являются мишенью для пенициллина, который ингибирует их синтез и препятствует формированию клеточной стенки. Количественное содержание пептидогли-
кана влияет на способность бактерий окрашиваться по Граму. Бактерии, имеющие значительную толщину муреинового слоя (90-95 %), стойко окрашиваются генцианвиолетом в сине-фиолетовый цвет и носят название грамположительных бактерий. Грамотрицательные бактерии с тонким слоем пептидогликана (5-10 %) в клеточной стенке после действия спирта утрачивают генцианвиолет и дополнительно окрашиваются фуксином в розовый цвет. Клеточные стенки у грамположительных и грамотрицательных прокариот резко различаются как по химическому составу (см. табл. 1.1), так и по ультраструктуре (см. рис. 1.5).
?
j 'V
Рис.
1.5. Схематическое изображение клеточной
стенки у грамположительных (а) и
грамотрицательных (б) прокариот: 1
- цитоплазматическая мембрана; 2 -
пептидогликан; 3 - периплазматиче- ское
пространство; 4 - наружная мембрана; 5
- ДНК
а 6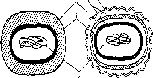
Химический
состав клеточных стенок грамположительных
и грамотрицательных прокариот
Таблица 1. 1
Компоненты клеточной стенки |
Грамположитель- ные прокариоты |
Грамотрицательные прокариоты |
|
внутренний слой (пептидоглика- новый) |
внешний слой (наружная клеточная мембрана) |
||
Пептидогликан |
+ |
+ |
- |
Тейхоевые кислоты |
+ |
- |
- |
Полисахариды |
+ |
- |
+ |
Окончание Табл. 1.1
Белки |
± |
— |
+ |
Липиды |
± |
— |
+ |
Липополисахариды |
— |
— |
+ |
Липопротеины |
— |
± |
+ |
Грамотрицательные прокариоты имеют наружную мембрану, в состав которой входят липиды (22 %), белки, полисахариды, липопро- теины.
Клеточная стенка у бактерий выполняет в основном формообразующую и защитную функции, обеспечивает ригидность, формирует капсулу, определяет способность клеток к адсорбции фагов.
Все бактерии, в зависимости от их отношения к окраске по Граму, делятся на грамположительные и грамотрицательные.
Методика окраски по Граму
На мазок кладут фильтровальную бумагу и наливают карболовый раствор генцианового фиолетового на 1-2 мин.
Снимают бумагу, сливают краситель и, не промывая мазок водой, наливают раствор Люголя на 1 мин.
Сливают раствор Люголя и обесцвечивают препарат в 96 %-м спирте в течение 30 сек.
Промывают водой.
Красят 1-2 мин водным раствором фуксина.
Промывают водой и высушивают.
В результате окраски грамположительные бактерии окрашиваются в фиолетовый цвет, грамотрицательные - в красный.
Причину различного отношения бактерий к окраске по Граму объясняют тем, что после обработки раствором Люголя образуется нерастворимый в спирте комплекс йода с генциановым фиолетовым. Этот комплекс у грамположительных бактерий, в связи со слабой проницаемостью их стенки, не может диффундировать, в то время как у грамот- рицательных - легко удаляется при промывании их этанолом, а затем водой.
Бактерии, полностью лишенные клеточной стенки, называются протопластами, они имеют шаровидную форму, обладают способностью к делению, дыханию, синтезу белков, нуклеиновых кислот, ферментов. Протопласты являются неустойчивыми структурами, очень
чувствительными к изменениям осмотического давления, механических воздействий и аэрации, не обладают способностью синтезировать составные части клеточной стенки, не подвергаются инфицированию вирусами бактерий (бактериофагами) и не обладают активной подвижностью.
Если под влиянием лизоцима и других факторов происходит частичное растворение клеточной стенки, то бактериальные клетки превращаются в сферические тела, получившие название сферопластов.
Под воздействием некоторых внешних факторов бактерии способны терять клеточную стенку, образуя L-формы (названы в честь института им. Д. Листера, где были впервые выделены); подобная трансформация может быть спонтанной (например, у хламидий) или индуцированной, например, под воздействием антибиотиков. Выделяют стабильные и нестабильные L-формы. Первые не способны к реверсии, а вторые реверсируют в исходные формы после удаления причинного фактора.
Цитоплазматическая мембрана
Цитоплазма бактериальной клетки ограничена от клеточной стенки тонкой полупроницаемой структурой толщиной 5—10 нм, называемой цитоплазматической мембраной (ЦПМ). ЦПМ состоит из двойного слоя фосфолипидов, пронизанных белковыми молекулами (рис. 1.6).
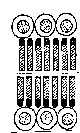
Рис.1.6. Строение плазматической мембраны Два слоя фосфолипидных молекул, обращенных гидрофобными полюсами друг к другу и покрытых двумя слоями молекул глобулярного белка.
С ЦПМ связаны многие ферменты и белки, участвующие в переносе питательных веществ, а также ферменты и переносчики электронов конечных стадий биологического окисления (дегидрогеназы, цито- хромная система, АТФ-аза). На ЦМП локализуются ферменты, катализирующие синтез пептидогликана, белков клеточной стенки, собствен
ных структур. Мембрана является также местом превращения энергии при фотосинтезе.
Периплазматическое пространство
Периплазматическое пространство (периплазма) представляет собой зону между клеточной стенкой и ЦПМ. Толщина периплазмы составляет около 10 нм, объем зависит от условий среды и прежде всего от осмотических свойств раствора. Периплазма может включать до 20 % всей находящейся в клетке воды, в ней локализуются некоторые ферменты (фосфатазы, пермеазы, нуклеазы и др.) и транспортные белки — переносчики соответствующих субстратов.
Цитоплазма
Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Та часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит растворимые РНК, ферменты, субстраты и продукты обмена веществ, обозначается как цитозоль. Другая часть цитоплазмы представлена различными структурными элементами: мезо- сомами, рибосомами, включениями, нуклеоидом, плазмидами.
Рис.
1.7. Рибосома (а) и ее субчастицы — большая
(б) и малая (в) Рибосомы
служат местом синтеза белка.
а б Е
Цитоплазматические
включения
Нередко
в цитоплазме бактерий обнаруживаются
различные включения, которые образуются
в процессе жизнедеятельности: капельки
нейтральных липидов, воска, серы,
гранулы гликогена, в-
гидроксимасляной кислоты (особенно у рода Bacillus). Гликоген и в- гидроксимасляная кислота служат для бактерий запасным источником энергии. У некоторых бактерий в цитоплазме находятся кристаллы белковой природы, обладающие ядовитым действием на насекомых.
Некоторые бактерии способны накапливать фосфорную кислоту в виде гранул полифосфата (зерна волютина, метахроматические зерна). Они играют роль фосфатных депо и выявляются в виде плотных образований в форме шара или эллипса, располагающихся в основном у полюсов клетки. Обычно на полюсах бывает по одной грануле.
Нуклеоид
Нуклеоид — ядерный аппарат бактерий. Представлен молекулой ДНК, соответствующей одной хромосоме. Она замкнута, располагается в ядерной вакуоле, не имеет ограничивающей от цитоплазмы мембраны.
С ДНК связано небольшое количество РНК и РНК-полимеразы. ДНК свернута вокруг центрального стержня, состоящего из РНК, и представляет собой высокоупорядоченную компактную структуру. Хромосомы большинства прокариот имеют молекулярную массу в пределах 1—3х109, константу седиментации 1300-2000 S. Молекула ДНК включает 1,6х10 нуклеотидных пар. Различия в генетическом аппарате прокариотических и эукариотических клеток обусловливают его название: у первых — нуклеоид (образование, подобное ядру), в отличие от ядра у вторых.
В нуклеоиде бактерий содержится основная наследственная информация, которая реализуется в синтезе специфических белковых молекул. С ДНК бактериальной клетки связаны системы репликации, репарации, транскрипции и трансляции.
Нуклеоид в прокариотической клетке может быть выявлен в окрашенных препаратах с помощью светового или фазово-контрастного микроскопа.
У многих бактерий в цитоплазме обнаружены внехромосомные генетические элементы — плазмиды. Они представляют собой замкнутые в кольца двухцепочечные ДНК, состоящие из 1500—40000 пар нук- леотидов и содержащие до 100 генов.
Капсула
Капсула — слизистый слой клеточной стенки бактерий, состоящий из полисахаридов или полипептидов. Микрокапсулу (толщиной менее 0,2 мкм) способны формировать большинство бактерий.
Жгутики
Жгутики выполняют роль органа движения, позволяющего бактериям передвигаться со скоростью 20—60 мкм/сек. Бактерии могут иметь один или несколько жгутиков, располагающихся по всей поверхности тела либо собранных в пучки у одного полюса, у разных полюсов. Толщина жгутиков в среднем составляет 10-30 нм, а длина достигает 10-20 мкм.
Основу жгутика составляет длинная спиральная нить (фибрилла), которая у поверхности клеточной стенки переходит в утолщенную изогнутую структуру - крюк и прикрепляется к базальной грануле, вмонтированной в клеточную стенку и ЦПМ (см. рис. 1.8).
Рис.
1.8. Схематическая модель базального
конца жгутика
Е. coli,
основанная на электронных
микрофотографиях выделенной органеллы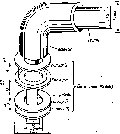
Базальные гранулы имеют диаметр около 40 нм и состоят из нескольких колец (одна пара - у грамположительных бактерий, четыре - у грамотрицательных прокариот). Удаление пептидогликанового слоя клеточной стенки ведет к потере способности бактерий к движению, хотя жгутики при этом остаются неповрежденными.
Жгутики почти полностью состоят из белка флагеллина с некоторым содержанием углеводов и РНК.
Споры
Некоторые бактерии в конце периода активного роста способны образовывать споры. Этому предшествует обеднение среды питательными веществами, изменение ее рН, накопление ядовитых продуктов метаболизма. Как правило, одна бактериальная клетка образует одну спору - локализация спор различна (центральная, терминальная, субтерминальная - рис. 1.9).
Рис. 1.9. Типичные формы спорообразующих клеток
Если размеры спор не превышают поперечного размера палочковидной бактерии, то последняя называется бациллой. Когда диаметр споры больше - бактерии имеют форму веретена и носят название кло- стридий.
По химическому составу различие спор от вегетативных клеток состоит лишь в количественном содержании химических соединений. Споры содержат меньше воды и больше липидов.
В состоянии споры микроорганизмы метаболически неактивны, выдерживают высокую температуру (140-150 °С) и воздействие химических дезинфицирующих веществ, длительно сохраняются в окружающей среде.
Попадая в питательную среду, споры прорастают в вегетативные клетки. Процесс прорастания спор включает три стадии: активации, начальной стадии и стадии роста. К активирующим агентам, нарушающим состояние покоя, относят повышенную температуру, кислую реакцию среды, механические повреждения и др. Спора начинает поглощать воду и с помощью гидролитических ферментов разрушает многие собственные структурные компоненты. После разрушения наружных слоев наступает период формирования вегетативной клетки с активацией биосинтеза, заканчивающейся делением клетки.