Глава 4. Генная инженерия
Генная инженерия - это раздел молекулярной генетики, связанный с целенаправленным созданием новых комбинаций генетического материала. Основа прикладной генетической инженерии - теория гена. Созданный генетический материал способен размножаться в клетке- хозяине и синтезировать конечные продукты обмена.
Генетическая инженерия возникла в 1972 г. в Станфордском университете США. Тогда лаборатория П. Берга получила первую реком- бинантную (гибридную) ДНК. Она соединяла в себе фрагменты ДНК фага лямбда, кишечной палочки и обезьяньего вируса SV40.
Генная инженерия - это направление исследований в молекулярной биологии и генетике, конечной целью которых является получение с помощью лабораторных приемов организмов с новыми, в том числе и не встречающимися в природе, комбинациями наследственных свойств. В основе генной инженерии лежит обусловленная последними достижениями молекулярной биологии и генетики возможность целенаправленного манипулирования с фрагментами нуклеиновых кислот. К этим достижениям следует отнести установление универсальности генетического кода, т. е. факта, что у всех живых организмов включение одних и тех же аминокислот в белковую молекулу кодируются одними и теми же последовательностями в цепи ДНК; успехи генетической энзимоло- гии, представившей в распоряжение исследователя набор ферментов, позволяющих получить в изолированном виде отдельные гены или фрагменты нуклеиновой кислоты, осуществлять in vitro синтез фрагментов нуклеиновых кислот, объединить в единое целое полученные фрагменты. Таким образом, изменение наследственных свойств организма с помощью генной инженерии сводится к конструированию из различных фрагментов нового генетического материала и введение его в рецепиентный организм, создания условий для его функционирования и стабильного исследования.
4.1. ИССЛЕДОВАНИЕ МОЛЕКУЛ ДНК
За последние годы методы исследования ДНК получили колоссальное развитие. К самым совершенным методам, с помощью которых изучается ДНК, относятся методы создания молекул ДНК путем соединения последовательностей, имеющих совершенно различное происхождение. Получаемый продукт называют рекомбинантной ДНК. Ре- комбинантная ДНК содержит ген (или гены) и вектор. Вектор - это фрагмент ДНК, обеспечивающий размножение гибридной ДНК и синтез конечных продуктов деятельности генетической системы - белков. Основные исследования выполнены на бактериях и вирусах, так как они являются одними из самых простых организмов, иначе их называют клетками «хозяина». Технология рекомбинантной ДНК позволяет получать генетические видоизмененные варианты целенаправленным и строго контролируемым путем.
Каким же образом гены высших организмов могут быть введены в бактериальные клетки? Наиболее распространенным методом генной инженерии является метод получения рекомбинантных ДНК, т. е. содержащих чужеродный ген, плазмид. Плазмиды представляют собой кольцевые двухцепочечные молекулы ДНК, состоящие из нескольких тысяч пар нуклеотидов. Каждая бактерия, помимо основной, не покидающей клетку молекулы ДНК (5 106 пар нуклеотидов), может содержать несколько различных плазмид, которыми она обменивается с другими бактериями. Плазмиды являются автономными генетическими, реплицирующими (т. е. размножающимися) в бактериальной клетке не в то же время, что основная молекула ДНК. Хотя на долю плазмид приходится лишь небольшая часть клеточной ДНК, именно они несут такие жизненно важные гены для бактерий, как гены лекарственной устойчивости. Разные плазмиды содержат различные гены устойчивости к антибактериальным препаратам. Простота устройства плазмид и легкость, с которой они «входят» и «выходят» из бактерий, используется генными инженерами для введения в клетки бактерий генов высших организмов.
Мощным инструментом генной инженерии являются открытые в 1974 г. ферменты - рестрикционные эндонуклеазы, или рестриктазы.
Рестрикция буквально означает «ограничение». Бактерии клетки вырабатывают рестриктазы для разрушения инородной (в первую очередь фаговой ДНК), что необходимо для ограничения вирусной инфекции. Рестриктазы «узнают» определенные последовательности нуклео- тидов в ДНК (так называемые сайты - участки узнавания) и вносят симметричные разрывы в цепях ДНК на равных растояниях от центра сайта. В результате на концах каждого фрагмента рестриктированных ДНК образуются короткие одноцепочечные «хвосты», которые называются «липкими концами».
Из разных видов бактерий выделено около пятисот различных ре- стриктаз, для которых описаны сайты рестрикции.
4.2. МЕТОДЫ ГЕННОЙ ИНЖЕНЕРИИ
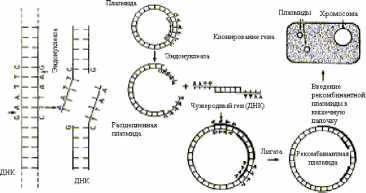
Для получения рекомбинантной плазмиды ДНК одна из плазмид расщепляется выбранной рестриктазой. Гены, которые нужно ввести в бактериальную клетку, выщепляют из ДНК хромосом человека с помощью той же рестриктазы, поэтому его «липкие концы» являются комплементарными нуклеотидным последовательностям на концах плазми- ды. Ферментом лигазой «сшивают» оба конца ДНК (гена и плазмиды), в результате получается рекомбинантная кольцевая плазмида, которую вводят в бактерию E. coli. Все потомки этой бактерии называются клоном и содержат в плазмидах чужеродный ген, способный вырабатывать белок, кодируемый этим геном. Весь процесс получения таких бактерий, называют клонированием. Он состоит из последовательных стадий (см. рис. 4.1):
Рестрикции - разрезания ДНК человека рестриктазой на множество различных фрагментов, но с одинаковыми «липкими» концами. Такие же концы получают при разрезании плазмидной ДНК той же ре- стриктазой.
Лигирования - включения фрагмента ДНК человека в плазмиды благодаря сшиванию «липких концов» ферментом лигазой.
Трансформации - введения рекомбинантных плазмид в бактериальные клетки, обработанные специальным образом - так, чтобы они на короткое время стали проницаемыми для макромолекул. Однако плазмиды проникают лишь в часть обработанных бактерий. Их разделяют, используя определенную питательную среду (например, раствор антибиотика). Каждая из трансформированных бактерий размножается и образует колонию из многих тысяч потомков - клон.
Скрининга - отбора среди клонов трансформированных бактерий тех, которые сохраняют плазмиды, несущие ген человека.
Не всегда удается точно вырезать нужный ген с помощью ре- стриктаз. Многие гены расщепляются этими ферментами на несколько частей или не содержат последовательностей, узнаваемых рестриктаза- ми. Поэтому в ряде случаев процесс клонирования начинают не с вырезания из хромосом случайных фрагментов ДНК, а с целенаправленного получения нужного гена. Для этого из клеток человека выделяют и- РНК, которая является транскрипционной копией этого гена, и с помощью фермента - обратной транскриптазы (ревертазы) - синтезируют комплементарную цепь ДНК, после чего и-РНК, служащая матрицей при синтезе ДНК, уничтожается РНК-азой - специальным ферментом, способным гидролизовать цепь РНК. Оставшаяся цепь ДНК служит матрицей для синтеза обратной транскриптазой (ДНК-полимераза) комплементарной второй цепи ДНК. Получаемая двойная спираль ДНК носит название к-ДНК (комплементарная ДНК), она соответствует гену, с которого была считана и-РНК. Такая к-ДНК встраивается в плазмиду, которая трансформирует бактерии, и получают клоны, содержащие только выбранные гены человека.
Рис.
4.1. Введение гена в плазмиду
Escherichia coli
и клонирование этого гена в клетках
кишечной палочки
Плазмида
E.coli расщепляется
рестрикционной эндонуклеазой в
специфическом участке в обеих цепях
ДНК, так что на концах расщепленной
плазмиды располагаются короткие
неспаренные последовательности
дезоксирибону- клеотидов (ТТАА или
ААТТ), т. е. по четыре нуклеотида, в
которых основания представлены тимином
и аденином.
Ген,
который нужно встроить в плазмиду,
выщепляют с помощью этой же рестриктазы,
так что его концы являются
комплементарными нуклеотидным
последовательностям на концах плазмиды
(ААТТ и ТТАА).
Обе
ДНК (гена и плазмиды) сшивают вместе с
помощью лигазы. Затем рекомбинантную
плазмиду вводят в клетку E.coli,которая,
размножаясь, образует клон, все клетки
которог содержат рекомбинантную
плазмиду, а поэтому и чужеродный ген.
Последний теперь клонирован в клетках
кишечной палочки и индуцирует в ней
синтез специфиче- скогобелка.