

Качественные реакции на пептиды и белки.
Биуретовая реакция (обнаружение в молекулах пептидов и белков пептидных связей)
В одну пробирку наливают 2 мл 1% раствора белка, в другую 2 мл 0.01% раствора глицина, а в третью – столько же дистиллированной воды,
Затем в каждую из них вносят по 2 мл 10% раствора NaOH и хорошо перемешивают.
Добавляют по 3 капли 2% раствора CuSO4. Пробирки встряхивают.
Отмечают появление окраски, объясняют различия окрашивания.
Оформление работы
К занятию:
Кратко законспектировать теоретический материал по лабораторной
работе.
Во время занятия:
Описать этапы лабораторной работы.
Зарисовать результаты хроматографического разделения аминокислот.
Описать результаты выполнения качественных реакций на аминокислоты.
Сделать выводы.
Методические указания к лабораторному практикуму по курсу
«Общая и экологическая биохимия»
Раздел «Белки»
Лабораторная работа № 2
Тема: |
Простые белки Физико-химические свойства белков |
Цель работы: |
|
Оборудование и материалы:
Термостат
Пипетки стекляные на 1 мл и 5 мл
Микропипетки автоматические
Цилиндры мерные на 250 мл и 100 мл
Колба емкостью 100 мл и 250 мл
Пробирки стекляные
Штативы для пробирок
Бумага фильтровальная
Индикаторная бумага универсальная
Сито нейлоновое
Воронки стекляные для фильтрации
Реактивы:
Яичный белок
Буферные растворы, рН 1.0, 3.7, 4.7, 5.7, 9.0
Гидроксид натрия (NaOH), 10% раствор
Хлорид натрия (NaCl), крист.
Хлорид натрия (NaCl), насыщенный раствор
Сульфат аммония ([NH4]2SO4), крист.
Сульфат аммония ([NH4]2SO4), насыщенный раствор
Сульфат магния (MgSO4), крист.
Сульфат меди (CuSO4), 2% раствор
Ацетат свинца (Pb(CH3COO)2), раствор
Желатин, порошок, 0.5% 1% раствор
Казеин, раствор
Уксусная кислота (CH3COOH), 1% раствор
Уксусная кислота (CH3COOH), 10% раствор
Азотная кислота (HNO3), конц., 5% раствор
Серная кислота (H2SO4), конц.
Соляная кислота (HCl), конц.
Трихлоруксусная кислота (ТХУ), 5% раствор
Спирт этиловый (C2H5OH), 96%
Ацетон
Фенол, насыщенный водный раствор
Формалин
Пикриновая кислота, насыщенный раствор
Таннин, 10% раствор
Гексацианоферриат калия (K3[Fe (CN)6]), 5% раствор
Вода дистиллированная
Теоретическая часть
Белки
Все белки являются высокомолекулярными полипептидами. Условную границу между крупными полипептидами и белками провести сложно. Обычно к белкам относят полипептиды с молекулярной массой, превышающей 8000-10000 дальтон. Белки бывают простыми и сложными. К простым белкам относят макромолекулы, состоящие только из аминокислот. Сложные белки включают неаминокислотные компоненты, такие как гем, производные витаминов, липиды, углеводы, атомы металлов и др.
Простые белки
Универсальной системы классификации белков не существует. Имеется лишь несколько общеупотребимых систем классификации, частично перекрывающихся между собой. Здесь мы рассмотрим основные принципы классификации белков, основанные на их растворимости, форме молекул, функциях, физических свойствах и особенностях трехмерной структуры.
Растворимость
Классификация белков, основанная на их растворимости, была введена в 1907-1908 годах и используется до сих пор (см. табл. 1).
Таблица 1
Растворимость наиболее известных типов белков
-
Тип белков
Характеристика
Альбумины
Растворимы в воде и солевых растворах.
По содержанию отдельных амино- кислот особенностей не имеют.
Глобулины
Слаборастворимы в воде, но хорошо растворимы в солевых растворах.
По содержанию отдельных амино-кислот особенностей не имеют.
Протамины
Растворимы в 70-80%-ном этаноле, но не растворимы в воде и в абсолютном этаноле.
Богаты аргинином.
Гистоны
Растворимы в солевых растворах. Богаты основными аминокисло-тами.
Склеропротеины
Нерастворимы в воде и солевых растворах.
Повышено содержание Gly, Ala, Pro.
Строго установленных границ между отдельными классами простых белков не существует. Например, четкое разграничение между альбуминами и глобулинами невозможно, если исходить только из их растворимости в воде и солевых растворах. Поэтому глобулины дополнительно подразделяют на псевдоглобулины, легко растворимые в воде, и эуглобулины, нерастворимые в воде, в отсутствие солей.
Форма молекул
Если исходить из оценки соотношения длины осей (продольной и поперечной), можно выделить два больших класса белков. У глобулярных белков это отношение составляет величину, меньшую 10, а в большинстве случаев не превышает 3-4. Такие белки характеризуются компактной укладкой полипептидных цепей. Примером служит инсулин, альбумины и глобулины плазмы крови, многие ферменты. Фибриллярные белки, у которых соотношение длины осей превышает 10, состоят из пучков полипептидных цепей, спирально навитых друг на друга и связанных между собой поперечными ковалентными или водородными связями. Примерами фибриллярных белков служат кератин, миозин, коллаген и фибрин.
Функции
Белки также классифицируют в соответствии с их биологическими функциями. В соответствии с этим принципом классификации, белки подраз-деляют на структурные, каталитические и транспортные (см. табл. 2).
Таблица 2
Биологические функции белков
-
Функция
Белки
Каталитическая
Ферменты
Сократительная
Актин, миозин
Регуляция работы генов
Гистоны, негистоновые ядерные белки
Гормональная
Инсулин
Защитная
Фибрин, иммуноглобулины, интерферон
Регуляторная
Кальмодулин
Структурная
Коллаген, эластин, кератины
Транспортная
Альбумины (преносят билирубин, жирные кислоты и т.д.), гемоглобин (кислород), липопротеины (различные липиды), транс-феррин (железо)
Энергетическая
Различные белки
Группу каталитических белков (ферментов), которая включает большинство различных типов белков, подразделяют на классы в соответствии с типом катализируемой ими реакции.
Физические свойства
Для ряда белков существуют специальные системы классификации, позволяющие устанавливать различия в пределах семейств сходных белков. Например, широко используются две и обсуждается третья система номенклатуры липопротеинов плазмы. По одной системе липопротеины классифицируют в соответствии с их поведением в электрическом или гравитационном поле. Так на основе электрофоретической подвижности при рН 8.6 различают 1-, 2-, - и -липопротеины. Вторая система классификации липопротеинов основана на их плотности в гидратированном состоянии. В этом случае различают хиломикроны, ЛПОНП (липопротеины очень низкой плотности), ЛПНП (липопротеины низкой плотности), ЛПВП (липопротеины высокой плотности), ЛПОВП (липопротеины очень высокой плотности). Возможен и третий тип классификации, основанный на первичной структуре апобелков. В соответствии с этой системой различают шесть классов липопротеинов плазмы крови, характеризующихся присутствием апобелков А, B, C, D, E и F, соответственно. Апобелки можно различать, используя иммунологические критерии.
Трехмерная структура
Белки можно разграничивать на основе их трехмерной структуры. Основой для такого принципа классификации белков служит структурное сходство или различие ряда белков, выявляемое, главным образом, с помощью рентгеновской кристаллографии.
Связи, принимающие участие в формировании структуры белка
Структура большинства белков стабилизируется двумя типами прочных связей (пептидных и дисульфидных) и тремя типами слабых взаимодействий (водородных, гидрофобных и электростатических).
Пептидная связь
В структурных формулах пептидов связь между карбонильным углеродом и атомом азота изображается как одинарная, однако на самом деле эта связь носит характер частично двойной связи. Поэтому свободное вращение вокруг нее невозможно, и все четыре атома пептидной связи (рис. 2.1) лежат в одной плоскости.
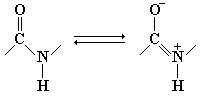
Рис. 2.1 Резонансная стабилизация пептидной связи придает ей характер частично двойной, чем объясняется заметная жесткость связи между атомами C и N.
Вращение же вокруг остальных связей полипептидного остова, наоборот, достаточно свободное. Эта полужесткость структуры имеет важное значение для формирования более высоких уровней структурной организации белковых молекул.
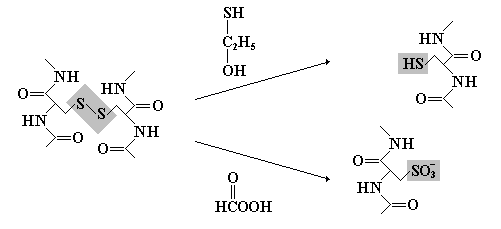
Межцепочечные и внутрицепочечные поперечные дисульфидные связи
Дисульфидные связи образуются между двумя остатками цистеина и «сшивают» два участка одной полипептидной цепи (или соседних цепей). Эта связь остается стабильной даже в тех условиях, при которых белки денатурируют. Обработка белка надмуравьиной кислотой (окисляющей S–S-связи) или -меркаптоэтанолом (восстанавливающим S–S-связи с регенерацией сульфгидрильных групп) приводит к разделению полипептидных цепей, связанных дисульфидными мостиками.
Стабилизация полипептидов межцепочечными и внутрицепочечными водородными связями
Водородные связи образуются: между боковыми цепями аминокислот; между атомами кислорода и водорода (связанного с атомом азотом), принадлежащими пептидным группам остова; между полярными остатками, расположенными на поверхности молекулы белка, и молекулами воды. Все эти взаимодействия играют важную роль в стабилизации вторичной и третичной структур белка.
Гидрофобные взаимодействия
Неполярные боковые цепи нейтральных аминокислот в белках имеют тенденцию к ассоциации. Стехиометрические соотношения при этом не соблюдаются. Тем не менее эти взаимодействиям играют важную роль в поддержании структуры белка.
Электростатические связи
Эти солевые связи возникают между разноименно заряженными группами боковых цепей аминокислот. Например, -аминогруппа лизина при физиологических рН несет заряд +1, а карбоксильная группа аспартата или глутамата в составе боковой цепи несет заряд 1. Следовательно, эти группы могут электростатически взаимодействовать, стабилизируя структуру белка.
Уровни структурной организации белка
Первичная структура
Под первичной структурой понимается последовательность аминокислот в полипептидной цепи (или цепях). Следовательно, первичная структура определяется тем, какие аминокислоты, сколько и в какой последовательности входят в состав полипептидной цепи.
Вторичная структура
Этот уровень структурной организации описывет стерические взаимосвязи между расположенными близко друг к другу аминокислотами. Вторичная структура может быть регулярной (-спираль, складчатый -слой) или не обнаруживать никаких признаков регулярности (неупорядоченная конформация), но во всех случаях в ее основе лежат водородные связи между карбонильным кислородом одного остатка аминокислоты и водородом, принадлежащим азоту пептидного остова другой аминокислоты.
Упорядоченные конформации полипептидов
-Спираль
Основными характеристиками -спирали являются следующие:
-Спираль стабилизируется водородными связями между атомом водорода, у атома азота пептидной группы, и карбонильным кислородом аминокислотного остатка, отстоящего от данной пептидной группы на четыре позиции вдоль цепи.
В образовании водородной связи участвуют все пептидные группы. Это обеспечивает максимальную стабильность -спирали.
В образование водородных связей вовлечены все атомы азота и кислорода пептидных групп, что в значительной мере снижает гидрофильность -спирализованных областей и увеличивает их гидрофобность.
-Спираль образуется самопроизвольно и является наиболее устойчивой конформацией полипептидной цепи, отвечающей минимуму свободной энергии.
Правосторонняя -спираль, обычно обнаруживаемая в белках, намного стабильнее левосторонней.
Стабильность -спирали в значительной степени зависит от аминокислотного состава соответствующего участка полипептидной цепи (табл. 3).
Таблица 3
Влияние различных аминокислот на формирование -спирали
-
Способствуют
Дестабилизируют
Препятствуют
Ala
Arg
Pro
Asn
Asp
Hyp
Cys
Glu
Gln
Gly
His
Lys
Leu
Ile
Met
Ser
Phe
Thr
Trp
Tyr
Val
Некоторые аминокислоты препятствуют образованию -спирали, и в месте их расположения непрерывность -спирали нарушается. К ним относятся пролин (атом азота пролина является частью жесткой кольцевой структуры, и вращение вокруг связи N - C становится невозможным), а также аминокислоты с заряженными или объемными R-группами, которые электростатически или механически препятствуют формированию -спирали.
Складчатый -слой
Альтернативная упорядоченная вторичная структура белка – складчатый -слой – была предложена Полингом и Кори. В то время как в -спирали полипептидная цепь находится в конденсированном состоянии, в складчатом -слое цепи почти полностью вытянуты. В тех случаях, когда соседние полипептидные цепи складчатого -слоя идут в противоположных направлениях (за положительное принимается направление от N- к С-концу), структуру называют антипараллельной. Когда соседние цепи идут в одном направлении, структуру -слоя называют параллельной.
Области складчатой -структуры присутствуют во многих белках, причем встречается как параллельная, так и антипараллельная формы. В формировании таких структур могут участвовать от двух до пяти соседних полипептидных цепей. Во многих белках одновременно присутствуют и -спирали, и складчатая -структура.
Неупорядоченная конформация (клубок)
Те участки белковой молекулы, которые не относятся ни к спирализованным, ни к складчатым структурам, обычно называют неупорядоченными. В такой конформации может находится значительная часть белковой молекулы. Нужно отметить, что с точки зрения биологической значимости неупорядоченные участки белка столь же важны, как -спираль и -структура.
Третичная структура
Общее расположение, взаимную укладку различных областей, доменов и отдельных аминокислотных остатков полипептидной цепи называют третичной структурой белка. Четкую границу между вторичной и третичной структурами провести сложно, однако, под третичной структурой понимают стерические взаимосвязи между аминокислотными остатками, далеко отстоящими друг от друга. Важнейшую роль в образовании третичной структуры играют гидрофобные взаимодействия между неполярными радикалами аминокислот, обращенными вглубь молекулы и взаимодействия обращенных наружу полярных групп с молекулами воды, которые окружают полипептид. Таким образом, степень «упакованности» молекулы белка, его конформация зависят от соотношения гидрофобных и гидрофильных аминокислот, а также от степени полярности растворителя.
Четвертичная структура
Белки, состоящие из двух и более полипептидных цепей, связанных между собой нековалентными связями обладают четвертичной (олигомерной) структурой. Олигомеры стабилизируются водородными связями и электростатическими взаимодействиями между остатками, находящимися на поверхности полипептидных цепей. Индивидуальные полипептидные цепи, составляющие олигомеры получили название протомеров, мономеров или субъединиц.
Многие олигомерные белки состоят из двух или четырех протомеров и называются, соответственно, димерами или тетрамерами. Однако довольно часто встречаются олигомеры, содержащие более четырех протомеров. Это особенно характерно для регуляторных белков. Олигомерные белки играют особую роль во внутриклеточной регуляции: их протомеры способны изменять взаимную ориентацию, что приводит к изменению свойств всего олигомера. Наиболее изученным примером олигомерного белка является гемоглобин.
Роль первичной структуры в формировании более высоких уровней структурной организации белков трудно переоценить. Действительно, вторичная и третичная структуры белков, формирующиеся самопроизвольно, определяются последовательностью аминокислот в полипептидных цепях. Эти процессы детерминируются химическими группами, соединенными с -углеродными атомами аминокислотных остатков. Таким образом, можно сказать, что не существует независимого генетического контроля за формированием уровней структурной организации белков выше первичного, поскольку первичная структура специфически определяет и вторичную, и третичную, и четвертичную структуры. Нативной конформацией белка, по-видимому, следует считать термодинамически наиболее устойчивую структуру в конкретных условиях.
Макромолекулярные белковые комплексы
Полифункциональные макромолекулярные комплексы образуются в результате ассоциации функционально разных белков, каждый из которых обладает всеми четырьмя уровнями структурной организации. Такое компактное расположение белков позволяет им более эффективно выполнять свои функции. Макромолекулярные комплексы функционируют в системе транспорта электронов, участвуют в биосинтезе жирных кислот, метаболизме пирувата и других синтетических и катаболических путях.