

72 Факторы эволюции: мутирование, миграция, естественный отбор, дрейф генов
Первые три фактора являются детерминистическими, т.е. результат их действия предсказуем. Последний же фактор (дрейф генов), наоборот, является стохастическим, т.е. его результаты полностью предсказать нельзя (или/или). Факторы эти находятся постоянно во взаимодействии.
Мутирование (МУ) – совокупность процессов, приводящих к появлению генетически отличных индивидов. МУ является единственным источником появления первичной изменчивости, поэтому значение его, как фактора эволюции очень велико.
Значительная часть особей популяции – носители вновь возникших мутаций. Постоянно идущий в природе мутационный процесс ведет к изменению в популяции частоты одного аллеля по отношению к другому. Хотя по каждому отдельному гену давление мутационного процесса не велико, при наличие же большого числа генов в организме оно оказывает заметное действие на генетическую структуру популяции (в сочетании с генетической комбинаторикой – множественность новых сочетаний аллеля).
В больших популяциях мутантный аллель может закрепиться только при условии значительного селективного преимущества перед исходным аллелем. Большая часть мутантных аллелей никак не закрепляется в большой панмиктической популяции. Спонтанный мутационный процесс статистичен и ненаправлен.
Эволюционное значение МУ определяется, прежде всего тем, что он постоянно поддерживает высокую степень гетерогенности природных популяций – основу для действия других факторов эволюции и прежде всего естественного отбора. Можно сказать, что мутационный процесс – это фактор-поставщик элементарного эволюционнго материала. Мутационный процесс ведет к возникновению части того резерва наследственной изменчивости, который определит в будущем возможность приспособления популяций к тем или иным изменениям условий среды.
Миграция – мощный детерминистический фактор, способствующий: снижению разницы в частотах аллелей между популяциями и уравновешиванию частот при отсутствии противодействующих сил.
Между смежными популяциями миграция более интенсивна, однако ее эффективность не высока, потому что популяции генетически близки. Часто миграция бывает эффективна между удаленными популяциями, так как эти популяции локально адаптированы.
Миграция – это основной фактор, обеспечивающий обратимость микроэволюции (дрейф и отбор дифференцируют популяцию). Процесс миграции способствует целостности вида и распространению по ареалу наиболее ценных в селективном отношении мутаций. С участием этого механизма регулируется численность особей в популяции и происходит заселение новых мест обитания. Миграция тесно связана с отбором.
Естественный отбор (ЕО) – дифференциальное воспроизводство генетически различающихся особей. ЕО является основным фактором в синтетической теории эволюции.
Предпосылки действия естественного отбора: наличие генетической изменчивости, проявляющейся фенотипически; разная вероятность воспроизводства разных генотипов. Вероятность воспроизводства генотипа есть приспособленность (адаптивная ценность). Она определяется соответствием фенотипа, определяемого данным генотипом к условиям окружающей среды. То есть ЕО всегда действует на уровне фенотипов.
ЕО всегда носит статистический характер.
2 аспекта ЕО: ЕО сохраняет прежнюю или создает новую композицию изменчивости в популяции; ЕО обуславливает направленную потерю изменчивости популяции и дифференцирует популяции.
ЕО формирует признаки и свойства путем интегрирования отдельных случайных уклонений с незначительной адаптивной ценностью. ЕО увеличивает общую приспособленность популяции. ЕО – относительная вероятность передать свои гены следующему поколению.
Виды отбора: стабилизирующий (поддержание и повышение устойчивости реализации в популяции среднего, ранее сложившегося значения признака или свойства), движущий (способствует сдвигу среднего значения признака или свойства), дизруптивный (благоприятствует более чем одному фенотипу и действует против средних промежуточных форм), частотозависимый отбор, сегрегационный.
Дрейф генов (ДГ)
Термин введен С.Райтом и Д.Д.Ромашовым, А.А.Калмагоровым, Н.П.Дубининым.
В отечественной генетике ДГ – Автоматические процессы.
ДГ – обусловленный случайными причинами процесс изменения частот генов, приводящий к потере изменчивости, а именно: к снижению гетерозиготности, фиксации (q=1) или элиминации (q=0) аллеля.
Эффективность ДГ зависит от: эффективной численности популяции (абсолютное число особей и СП, динамика численности, системы скрещивания, подразделенность популяции, характер вклада генотипа в фонд гамет определяют эффективную численность популяции). ДГ также приводит к дифференциации смежных популяций, к выщеплению аллелей в гомозиготах с негативными эффектами, повышению вероятности формирования коадаптивных генов, к генетическому преобразованию популяции без генетического груза.
73. Естественный отбор – это один из факторов микроэволюции. Является направляющим фактором динамики генетического состава популяции.
Выделяют следующие типы естественного отбора:
• Стабилизирующий благоприятен тем организмам, чьи признаки ближе к среднему. Он всегда способствует сохранению полиморфизма. Отбор в пользу гетерозигот. Уменьшение крайних вариантов.
Д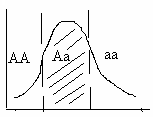 ля
того чтобы существовать, популяция
вынуждена постоянно приспосабливаться
к условиям
своего местообитания. На протяжении
всей истории
жизни популяции естественный отбор
стремится сохранить
те генотипы, которые приводят к образованию
фенотипов, наиболее хорошо соответствующих
комплексу
внешних условий, преобладающих в данное
время. Если в течение нескольких поколений
условия
существования остаются неизменными,
то популяция
имеет возможность достичь высокого
уровня
приспособленности
ля
того чтобы существовать, популяция
вынуждена постоянно приспосабливаться
к условиям
своего местообитания. На протяжении
всей истории
жизни популяции естественный отбор
стремится сохранить
те генотипы, которые приводят к образованию
фенотипов, наиболее хорошо соответствующих
комплексу
внешних условий, преобладающих в данное
время. Если в течение нескольких поколений
условия
существования остаются неизменными,
то популяция
имеет возможность достичь высокого
уровня
приспособленности
Это значит, что некоторая группа хорошо приспособленных генотипов (адаптивная норма) сохраняется из поколения в поколение, тогда как формы, отклоняющиеся от нормы, элиминируются в каждом цикле отбора. Отбор не прекращается потому, что эти слабо приспособленные формы постоянно возникают за счет одного или нескольких источников изменчивости — мутационного процесса, миграции, рекомбинации и выщепления.
Эти наблюдения показали, что нередко именно эти формы, соответствующие крайним значениям метрического признака, наиболее слабо приспособлены.
В качестве классического примера можно привести работу Бампуса, более 60 лет назад изучавшего дифференциальное выживание в группе воробьев (Passer domestica), которых он подобрал в жестокую пургу и принес в лабораторию. Из всех этих беспомощных и полузамерзших птиц выжила лишь половина. Проведя измерения некоторых морфологических признаков у живых и мертвых птиц, Бампус пришел к выводу, что у выживших воробьев признаки близки к средним для вида значениям (адаптивная норма), тогда как среди погибших птиц эти признаки сильно варьировали. Позднее Керн и Пенроуз установили связь между смертностью и весом новорожденных. Оказалось, что наибольшая смертность в первые 28 дней жизни наблюдалась среди детей, вес которых при рождении был значительно больше или значительно меньше среднего . Стабилизирующий отбор иногда приводит к некоторому снижению изменчивости признака, причем это снижение не сопровождается изменением среднего значения признак
Направленный Оптимальную приспособленность имеет один из крайних фенотипов. Отбор способствует сохранению признаков, благоприятных при существующих условиях, и создаваемая таким образом приспособленность отнюдь не гарантирует каких-то преимуществ ее носителям в будущем. Чем дольше популяция находится в условиях стабильного местообитания, тем выше ее шансы достигнуть приспособленности и сохранить ее впредь. Если популяция оказывается в новой среде, то следует ожидать, что вместо стабилизации изменчивости будет наблюдаться прямо противоположная картина — резкое ускорение эволюционного процесса. При этом может оказаться, что наиболее редкие варианты, прежде сравнительно плохо приспособленные, будут сохраняться отбором и, следовательно, фенотипический оптимум последующих поколений будет отличаться от фенотипической нормы, характерной для предков.
Отбор, при котором оптимальной приспособленностью обладают фенотипы, расположенные не в середине, а ближе к одному из концов кривой распределения частот фенотипов, называется направленным
Дизруптивный Третий тип отбора можно рассматривать как обратный по отношению к стабилизирующему отбору в том смысле, что при этом множество различных сил действует на различные формы одной популяции. Отбор, благоприятствующий более чем одному фенотипическому оптимуму, т. е. совершенно различным генотипам, и действующий против промежуточных форм, называется дизруптивным, или разрывающим.
Ожидаемый ответ популяции на дизруптивный отбор— установление прерывистости в изменчивости. В итоге дизруптивный отбор может привести;
1) к полиморфизму — состоянию, при котором каждый оптимум представлен различной морфой;
2) к дивергенции и изоляции — состояниям, при которых оптимумы представлены особями, образующими локально адаптированные «субпопуляции».
В любом случае конечный результат сводится к направляющему действию, т. е. каждому классу отбираемых вариантов свойственно свое направление изменений, свой оптимум, к которому и стремится структура группы
По-видимому, в природе некоторые виды полиморфизма обусловлены именно дизруптивным отбором. В качестве примера можно привести множество мимикрирующих форм бабочек у многих географических рас африканского парусника Papilio dardanus. Каждая из нескольких обычных для данной местности форм сходна с каким-либо обитающим в той же местности несъедобным видом бабочек. Это сходство сравнительно съедобных форм с несъедобными и предохраняет первых от уничтожения птицами. Как показывают многочисленные данные, такой тип мимикрии особенно эффективен в том случае, когда численность подражающих особей значительно меньше численности особей, служащих моделью. Это объясняется тем, что хищникам необходимо многократно убеждаться в несъедобности модели, для того чтобы научиться избегать ее. Если же несъедобная модель встречается редко, то мимикрирующая форма не сможет сохраниться в течение сколько-нибудь долгого времени, поскольку отвращение у хищников к внешнему виду модели будет выражено слабо. Отсюда был сделан вывод, что значительно благоприятнее для таких видов, как P. dardanus, подражать не одной, а нескольким несъедобным формам. По-видимому, полиморфизм у этого вида возник в результате дизруптивного отбора — различные морфы (оптимумы) функционально связаны, поскольку успех каждой из них зависит от наличия других морф.