

Радиобиология / Моссэ И. Б., Морозик П. М. Генетические эффекты ионизирующей радиации
.pdfполовина окажется с вырезками на крыльях и белыми глазами. Очевидно, у таких самок рецессивный ген W проявился вследствие того, что доминант- ная аллель w + в гомологичной хромосоме отсутствует. Другие два рецессив- ных гена – у и f у этих самок не проявляются, потому что их действие пода- вляется сохранившимися доминантными аллелями у+ и f +. Значит, выпавший участок Х-хромосомы у мух Notch относится только к району локуса W. Поэто-
му генотип этих мух можно представить следующим образом:
y+ f + . ywf
Такое явление, когда рецессивный ген проявляется в гетерозиготном со- стоянии, в случае выпадения доминантной аллели называют ложным доми- нированием, или псевдодоминированием. Рецессивный ген при этом находит-
ся в гемизиготном состоянии. Он может проявляться также у диплоидных организмов, у которых нацело отсутствует одна из гомологичных хромосом.
Можно привести также пример наследования нехватки в аутосоме у до-
машней мыши. У мыши известна рецессивная мутация, обусловливающая круговые «вальсирующие» движения. Если скрещивается самка, гомозиготная по этому гену W, с нормальным самцом, несущим доминантный ген W+ той же аллельной пары, то в первом поколении все потомство оказывается нормаль- ным. Но иногда в F1 появляются «вальсирующие» особи. Когда проверили ци- тологически соматические клетки этих мышей, то оказалось, что одна из хро- мосом имела делецию. Выпадение доминантного гена W+ позволило проявить- ся рецессивному гену «вальсирования», находившемуся у гибрида в гемизи-
готном состоянии.
Однако некоторые гены, находясь в гемизиготном состоянии у самок и самцов дрозофилы, проявляются по-разному. Как было показано в опытах М. Е. Лобашева в 1935 г., при скрещивании самцов дрозофилы с геном w a (абрикосовый цвет глаз) или w co (коралловый) с самками Notch, имеющими в одной хромосоме делецию в районе локуса white, а в целой хромосоме одну из данных аллелей, в потомстве получаются два класса самок, различающих-
ся по окраске глаз, а также по наличию или отсутствию вырезки на крыле: у самок без вырезки крыла окраска глаз сходна с окраской глаз самца, а у са- мок с вырезкой на крыле глаза значительно светлее, чем у самцов. Таким об-
разом, гемизиготное состояние гена у нормального самца и самки с делецией проявляется различно: одна доза гена у самца не равна по действию одной дозе того же гена у самки. Гемизиготное состояние указанных аллелей у нор-
мального самца и двойная доза у нормальной самки (без делеции) проявляется одинаковым образом.
Другие аллели этой серии, а именно We и WB (соответственно эозиновый
и кровяной), обнаруживают иной характер действия (рис. 1.6).
Эти гены в гемизиготном состоянии у самок и самцов проявляются одина-
ково, а в двойной дозе у нормальных самок дают более темную окраску. Здесь
40
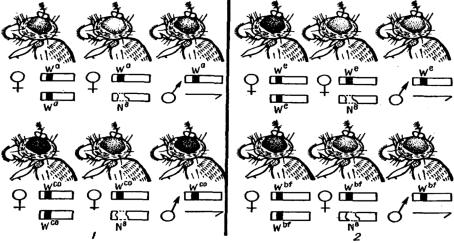
Рис. 1.6. Фенотипическое проявление множественных аллелей локуса white в гомо- и гемизи- готном состояниях у Drosophila: 1 – с компенсаторным эффектом у самцов; 2 – без компенса- торного эффекта у самцов; N8 – делеция Notch [1]
мы не будем касаться причин, вызывающих описанные различия, так как они далеко не выяснены, однако данные результаты иллюстрируют два очень важных положения. Во-первых, выпадение участка, в одной из гомологичных хромосом дает возможность проявиться рецессивным генам в гемизиготном состоянии в другом гомологе; во-вторых, аллели в гемизиготном состоянии у самок и самцов могут проявиться неодинаково.
На основе использования гетерозиготных делеций у диплоидных организ-
мов по схемам скрещиваний, аналогичным рассмотренной, можно обнаружить рецессивные мутации тех генов, которые противостоят нехватке в одном из двух гомологов. Так, например, у дрозофилы установлено около 130 делеций различной длины типа Notch.
На дрозофиле был разработан также специальный метод так называемых перекрывающихся делеций для локализации генов в хромосоме. Получая ряд независимых делеций разной длины в одном районе хромосомы и комбинируя их путем скрещивания с соответствующими линиями мух, можно цитологи-
чески локализовать место гена в хромосоме. На рис. 1.7 показан пример картирования генов на основе получения перекрывающихся делеций в районе white – Notch Х-хромосомы дрозофилы. Для цитологического картирования генов также использовались нехватки, возникающие в хромосомах кукурузы. Принципиально этот же метод был использован для внутригенного картиро-
вания цистронов А и В локуса rll у бактериофага Т4.
Как же осуществляется конъюгация гомологичных хромосом в мейозе в случае нехватки участка в одной из гомологичных хромосом? В мейотиче-
ских хромосомах это трудно наблюдать. Лишь когда утрачивается достаточно
41
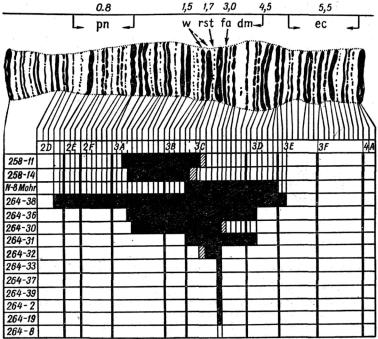
Рис. 1.7. Генетическая карта white – Notch-района Х-хромосомы Drosophila melanogaster вместе с диаграммой, указывающей протяженность четырнадцати делеций, которые дают фенотип white – Notch: черные участки – определенные границы делеций, заштрихованные участки – не определенные делеций; цифрами обозначены диски гигантской хромосомы, буквами – группы дисков; цифры, расположенные вертикально, – номера мутантов с соответствующими делециями; обозначение генов: рN – prune, rst – roughest, fa – faset, dm – diminutive, ec – echinus,
w – white [1]
большой участок хромосомы, такие нехватки можно обнаружить в пахитен-
ной стадии, так как на этой стадии хромосомы выглядят в виде тонких нитей с хромомерами.
Но наиболее отчетливо нехватки можно видеть в гигантских хромосомах. Го- мологичные хромосомы слюнных желез дрозофилы в норме конъюгируют до- вольно плотно (соматическая конъюгация). При этом идентичные диски оказыва-
ются тесно прилежащими друг к другу. Но в случае гетерозиготного состояния по нехватке внутреннего участка одной из хромосом нормальная конъюгация в этом районе нарушается. Диски такого участка нормальной хромосомы, не имея себе партнеров в другой хромосоме, образуют петлю, в то время как все осталь-
ные гомологичные диски в обеих хромосомах тесно прилегают друг к другу (рис. 1.8). Силы взаимного притяжения дисков в гомологичных хромосомах оста- ются неизвестными, но сам факт очевиден. Предполагают, что такой же тип конъ-
югации хромосом может иметь место и в профазе мейоза.
Дупликации. Прямой противоположностью нехваткам является умноже-
ние отдельных участков хромосом. Одновременно с возникновением нехватки
42
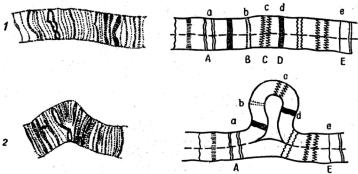
Рис. 1.8. Синапсис гомологичных хромосом при наличии нехватки в одной из них: 1 – нор-
мальные хромосомы; 2 – в одной из хромосом имеется нехватка по локусам BCD [1]
вследствие потери участка внутри одной хромосомы внутри другой гомоло-
гичной хромосомы могут накапливаться идентичные участки, несущие одни и те же гены. Явление умножения одного и того же участка хромосомы, со- держащего одни и те же гены, вызывающее фенотипическое изменение при-
знаков, называют повторением, или дупликацией.
Если в норме в каждой хромосоме ген представлен одной дозой, то при уд- воении или утроении несущего его участка доза гена соответственно увели- чится в 2 или 3 раза. Например, если гены в нормальной хромосоме располо-
жены в порядке ABC, то при дупликациях одного из генов могут возникнуть состояния АВВС или АВВВС и т. д. По-видимому, более частым случаем ду- пликации является повторение идентичных участков хромосомы, инкрусти-
рованных последовательно, например, ABC ABC ABC. Такой тип умножения идентичных наборов генов называют дупликацией «гуськом». Он обнаружен у домашней мыши, нейроспоры, аспергилла, кукурузы и др. Прекрасным при- мером накопления идентичных участков хромосомы является изменение фе- нотипического проявления формы глаз типа Ваr у дрозофилы. И. А. Рапопор-
ту удалось добиться умножения этого локуса в одной и той же хромосоме до 8 раз. Сходный факт дупликации был найден С. И. Алиханяном у дрозофилы для доминантной мутации Hairy wing (волосатые крылья). Это явление было подтверждено позднее американским генетиком М. Демерецем. По генетиче-
ским и цитологическим данным В. Льюиса, мутация asteroid, находящаяся в одном из концов второй хромосомы дрозофилы, является удвоением одного и того же участка хромосомы – локуса Star. В настоящее время накоплены до- полнительные сведения о том, что многие гены, принимавшиеся ранее за точ-
ковые мутации, оказались дупликациями: white, scute, acheate, vermilion и др. Дуплицированные участки могут находиться не только в соседних участ-
ках хромосомы, как это наблюдается в случае возникновения эффектов Ваr и asteroid, но и в других районах той же хромосомы (перемещенные дуплика- ции). В силу этого идентичные участки могут быть распределены по всей хро-
мосоме, а в некоторых случаях перемещены в другие хромосомы.
43
При дупликации двух идентичных генов, сходных по характеру действия и оказавшихся в разных группах сцепления при скрещивании, будет наблю- даться расщепление 15:1, т. е. такое же, как в случае дигибридного расщепле-
ния полимерных генов.
К дупликациям можно отнести также и явление умножения числа генов при увеличении числа гомологичных хромосом в наборе. В этом случае все признаки, определяемые генами, находящимися в данной группе сцепления, изменяются соответственно увеличению дозы гена. По-видимому, умножение идентичных участков является широко распространенным явлением в эволюции хромосом. Некоторые авторы полагают, что в основе усложнения гена как единицы наслед-
ственности лежит процесс умножения мелких идентичных участков хромосом. Инверсии. Инверсией называют изменение порядка расположения генов
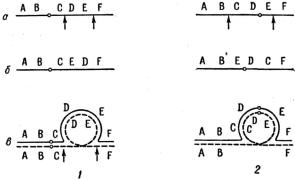
в хромосоме, вызванное перевертыванием на 180° большого или маленького участка внутри хромосомы. Если нормальную последовательность участков с содержащимися в них генами представить в алфавитном порядке АВСD, то
при инверсии этот порядок может измениться и стать АСВD (рис. 1.9).
Для образования инверсии внутри хромосомы необходим разрыв в двух точках, и только при этом условии участок может прикрепиться противопо-
ложными концами, перевернувшись на 180°.
Перевертывание концевого участка хромосомы обычно не встречается, вероятно, потому, что конец хромосомы – теломера не обладает свойством воссоединяться с разорванным концом хромосомы. Напротив, разорванным (открытым) концам хромосом свойственна высокая способность к воссоеди- нению. Инверсии часто связаны с рецессивным летальным эффектом, поэто- му они не сохраняются в гомозиготном состоянии, и их обычно не обнаружи- вают в гетерозиготе. Однако встречаются и такие инверсии, которые не связа-
ны с летальным эффектом.
Рис. 1.9. Типы инверсий: 1 – парацентрическая инверсия; 2 – перицентрическая инверсия; а – местоположение инвертированного участка по отношению к центромере; б – хромосома, содержащая инвертированный участок; в – образование кольца при синапсисе гомологичных хромосом, одна из которых содержит инверсию. Стрелки указывают на инвертированные районы [1]
44
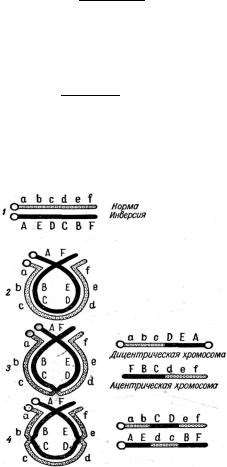
Существуют два типа инверсий – парацентрические и перицентрические. В случае парацентрической инверсии в хромосоме АВ○CDEF (где ○ означает центромеру) расположение генов может быть, например, AB○CEDF. В случае перицентрической инверсии последовательность генов ABCD○EF изменится на ABE○DCF (рис. 1.9).
При гомозиготной инверсии кроссинговер осуществляется нормально, при гетерозиготной кроссинговер подавляется полностью или частично. Так, например, в гетерозиготной парацентрической инверсии хотя и происходит конъюгация хромосом, как это показано на рис. 1.10, однако кроссоверных га- мет при одинарном кроссинговере не обнаруживается, и это создает впечатле-
ние подавления кроссинговера:
abcdef
AEDCBF .
Всилу нарушения нормального образования продуктов мейоза при скре-
щивании гетерозиготной по инверсии самки дрозофилы с самцом, несущим только рецессивные гены в гомозиготном состоянии
abcdef abcdef
AEDCBF abcdef
в потомстве не будет возникать кроссоверных особей по данным генам. На рис. 1.10, 3 можно видеть, что если происходит одинарный перекрест между хроматидами, то в результате образуются две необычные хромосомы: одна из них без центромеры, а другая – с двумя центромерами.
Рис. 1.10. Синапсис и перекрест в случае гетерозиготной инверсии в телоцентрической хромо-
соме и продукты мейоза: 1 – нормальная (abcdef) и инвертированная (AEDCBF) хромосомы до синапсиса; 2 – синапсис; 3 – одинарный перекрест и продукты мейоза (дицентрическая в ацентрическая хромосомы); 4 – двойной перекрест и продукты мейоза; светлым кружком обозначены центромеры [1]
45
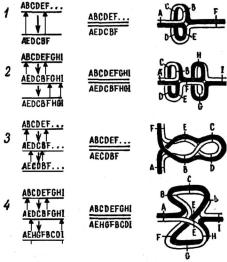
Последняя в анафазе при полярном расхождении центромер образует хромосомный «мостик», т. е. полярно растянутые хромосомы (рис. 1.10, 3). Мо- стик может разорваться в любом месте, в результате чего гаметы получат хро-
мосомы с более или менее значительными нехватками по одним участкам
ис дупликациями по другим и поэтому окажутся нежизнеспособными. Нормальные жизнеспособные гаметы могут образовываться только за счет
хроматид, не вступивших в перекрест. Кроссоверные гаметы оказываются не-
жизнеспособными, и создается впечатление об отсутствии или подавлении кроссинговера у гетерозигот.
При длинной инверсии, затрагивающей большой район хромосомы (на- пример, в линии СlВ), иногда может обнаруживаться кроссинговер внутри ин-
вертированного участка. В подобном случае кроссоверы возникают за счет двойного перекреста (рис. 1.10, 4), в гетерозиготных инверсиях по обе сторо-
ны от разрывов кроссинговер подавляется в силу отсутствия в этих районах конъюгации хроматид.
Влитературе инверсию как подавителя кроссинговера принято обозна- чать буквой С (начальная буква английского слова crossingover). Это обозна-
чение сохранилось с тех пор, когда подавление кроссинговера приписывалось особому гену в хромосоме.
Вотличие от парацентрических в перицентрических инверсиях ни оди-
ночный, ни двойной кроссинговер не дает ацентрических и дицентрических хроматид, а, следовательно, мостов и фрагментов в мейозе. В результате крос-
Рис. 1.11. Возможные типы |
инверсий: |
1 – одиночная инверсия; 2 |
– инверсия |
«гуськом»; 3 – полностью перекрывающаяся инверсия; 4 – частично перекрывающаяся инверсия [1]
синговера внутри перицентрических инверсий образуются хроматиды с дупликациями и нехватками. Пыльца и яйцеклетки высших растений, имеющие ука- занные хроматиды, оказываются не-
жизнеспособными. Перицентрические инверсии могут изменять место цен-
тромеры в группе сцепления. Инверсию можно обнаружить так-
же цитологически в гигантских хромосомах или на пахитенной стадии мейоза. Для того чтобы осуществилась конъюгация хромосомы, имеющей ин-
вертированный участок, с нормальной хромосомой у особи, гетерозиготной по инверсии, эти гомологичные хромосомы должны образовать петлю (рис. 1.11, 1). При этом гены точь-в-точь пригоняют-
ся друг к другу. Следовательно, между гомологичными локусами хромосом существует взаимное притяжение огром-
46
ной силы. Это очень важное явление для понимания физических и химиче- ских причин конъюгации гомологичных хромосом в профазе мейоза и в по-
литенных хромосомах.
Наряду с простыми одинарными инверсиями в хромосомах встречаются комплексы инверсий, которые могут состоять из нескольких одинарных неперекрывающихся инверсий, называемых иногда инверсиями «гуськом» (рис. 1.11, 2), или сложными инверсиями. В последние включаются полностью или частично перекрывающиеся двойные инверсии (рис. 1.11, 3, 4).
Итак, в генетическом отношении инверсии характеризуются следующими свойствами:
1)в гомозиготных инверсиях, если они не связаны с летальным эффектом, кроссинговер протекает нормально. Причина этого очевидна, поскольку та- кие гомологичные хромосомы в профазе мейоза могут свободно конъюгиро-
вать и обмениваться идентичными участками;
2)в гетерозиготных инверсиях, возникающих в двуплечих хромосомах, кроссинговер подавляется только в том плече хромосомы, в котором имеется инверсия, в другом плече подавления перекреста не обнаруживается.
Однако следует иметь в виду, что не всякое подавление кроссинговера может быть отнесено за счет действия инверсии. Существуют такие генные мутации, которые могут препятствовать нормальному синапсису хромосом
впрофазе мейоза (асинаптические гены), как это установлено у кукурузы, ржи, хлопчатника, дурмана, ячменя, а также у дрозофилы.
Инверсии встречаются в природных популяциях животных и растений, а также могут быть получены в эксперименте, особенно под влиянием иони-
зирующих излучений и ряда химических веществ. Генетики полагают, что инверсии имеют существенное значение для дивергенции видов. Так, рядом исследований установлено, что отдельные расы внутри вида, например, у Dro sophila pseudoobscura, могут различаться по инверсиям. Близкие виды дрозо-
филы в сходных хромосомах могут иметь обратный порядок расположения генов. Н. Н. Соколов провел цитогенетическое сравнение гигантских хромо- сом у двух видов дрозофилы (D. virilis и D. littoralis) и у их гибридов, в резуль- тате чего выяснилось, что эти два вида различаются между собой шестью ин- версиями и несколькими участками, не способными к нормальной соматиче-
ской конъюгации.
Экспериментально полученные инверсии часто используются в методиче-
ских целях для создания линий с подавленным кроссинговером. В п. 1.1 мы приводили примеры использования таких линий (СlВ и М-5) для учета ле- тальных мутаций, а также использования инверсий для локализации разры-
вов и генов.
Инсерции. Инсерцией называется перестановка участков внутри одной хромосомы. В результате перемещения генов из одного района в другой той же хромосомы первоначальные свойства генов могут либо изменяться, либо сохраняться. Эффект зависит от того, в каком новом сочетании они окажутся.
47
Инсерции изменяют порядок расположения генов в группе сцепления и конъ-
югацию хромосом в мейозе, тем самым уменьшая возможность рекомбинации генов.
1.2.2. Межхромосомные перестройки
Кроме рассмотренных внутрихромосомных перестроек существуют изменения, затрагивающие одновременно две или более негомологичные хро- мосомы. К таким перестройкам относятся транслокации, т. е. обмен участка-
ми между негомологичными хромосомами. При транслокациях изменяются группы сцепления генов.
Впервые явление обмена участками между негомологичными хромосома-
ми обнаружено Дж. Беллингом в 1915 г. Он наблюдал 50 %-ную стерильность пыльцы и 50 %-ную дегенерацию зародышевых мешков при определенной си- стеме скрещиваний – бархатных бобов Stizolobium deerigianum. В 1925 г. ука- занная полустерильность найдена ученым также у дурмана (Datura) и объяс- нена им как результат сегментных перестроек между негомологичными хро-
мосомами. У дрозофилы транслокация впервые описана Шерном в 1926 г., она касалась переноса участка Y- на Х-хромосому. Вскоре транслокации были
найдены также у кукурузы по полустерильности початков и пыльцы.
Для того чтобы произошла любая хромосомная перестройка, необходимо осуществление двух процессов: 1) разрыв хромосом и 2) воссоединение ото- рвавшихся кусков-сегментов с той же самой хромосомой (внутрихромосом- ные перестройки) или соединение с негомологичной хромосомой (межхромо-
сомные перестройки, или транслокации).
Допустим, что в норме одна пара хромосом несет гены
ABCDABCD ,
а другая пара – гены
EFGHEFGH .
При одновременном разрыве в двух негомологичных хромосомах оторвавшиеся сегменты взаимно обменяются местами, например:
ABGHABCD и EFGHEFCD .
При этом обменявшиеся участки могут быть равной или неравной длины и включать равное или неравное количество генов. Такой тип обмена называ- ют взаимной, или реципрокной транслокацией. Раньше считалось, что суще- ствуют и нереципрокные транслокации, однако в последнее время существо-
вание их подвергнуто сомнению.
При реципрокной транслокации может происходить объединение двух центромерных участков. В таком случае одна транслоцированная хромосома
48
окажется с двумя центромерами (дицентрическая), а два других сегмента ока-
жутся бесцентромерными и потеряются при делении клетки.
Главной особенностью хромосомных перестроек типа транслокаций явля-
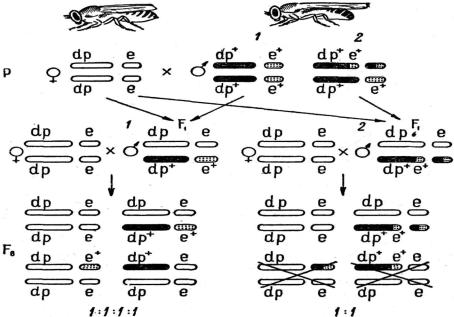
ется изменение групп сцепления. Перемещенные гены войдут в новые группы сцепления и тем самым нарушится сложившаяся система генотипа – на этом основаны генетические методы обнаружения транслокаций. При гибридоло- гическом анализе их устанавливают по изменению групп сцепления и нару-
шению генного баланса в зиготе (рис. 1.12).
Рассмотрим примеры обнаружения транслокаций у дрозофилы. Для скре- щивания берут линию дрозофил, маркированную двумя парами любых ре-
цессивных генов. Например, одна пара генов – укороченные крылья (dumpy) – находится во II группе сцепления, а другая – темно-серое тело (ebony) – в III.
С гомозиготной по этим генам самкой скрещивается нормальный самец
dp e dp+ e+ , dp e dp+ e+
половые клетки которого анализируются на наличие в них взаимной трансло- кации между II и парами хромосом.
Из F1 берут самцов, гетерозиготных по указанным генам, и индивидуаль-
но скрещивают их с самками из линии с двумя парами рецессивных генов (анализирующее скрещивание):
Рис. 1.12. Схема генетического анализа транслокаций: 1 – результаты скрещивания в норме; 2 – результаты скрещивания при наличии транслокаций; dp+ – нормальные крылья; dp – укоро-
ченные крылья; е+ – серое тело, е – темное тело [1]
49