

Радиобиология / Моссэ И. Б., Морозик П. М. Генетические эффекты ионизирующей радиации
.pdfвызванного воздействием облучения альфачастицами, до сих пор остается неясным. Тем не менее предполагают, что щелевидные сое-
динения между клетками участвуют в его распространении после действия низких доз облучения альфа-частицами.
«Байстэндер» эффект, вызванный воздей ствием бетачастиц. «Байстэндер» эффект,
вызванный воздействием бета-частиц, был продемонстрирован в многоклеточной моде-
ли с помощью группы клеток, помеченных тритиированным тимидином ([3H]dThd) [352, 363]. Малая область распространения бета-час-
тиц 3H вызывает лишь поражение меченых клеток, а немеченые клетки не повергаются воздействию излучения (рис. 4.7).
Однако в результате немеченые клетки также оказываются пораженными. Щелевидные соединения между клетками вовлекаются в байстэндер эффект, который имеет место до тех пор, пока линдан (гамма-изомер гексахлороци- клогексана), ингибитор межклеточных процессов, осуществляемых через ще-
левидные соединения, не проявит защитный эффект.
«Байстэндер» эффект, вызванный воздействием гаммалучей. К. Мазер-
сил и К. Сеймур разработали способ исследования «байстэндер» эффекта, индуцированного гамма-лучами [351]. Для этого использовали облучение клеточных культур эпителиальных клеток человека гамма-лучами. После об- лучения культуральную среду пропускали через специальные микропори-
стые фильтры для удаления клеток. Отфильтрованную питательную среду переносили к интактным клеткам (рис. 4.8).
Было показано, что культуральная среда от облученных гамма-лучами клеток повреждает необлученные клетки – в них наблюдалось снижение жиз-
Рис. 4.8. Способ индукции РИБЭ гамма-излучением с последующим переносом культураль-
ной среды от облученных клеток интактным
180
неспособности и колонеобразующей активности, а также возрастание частоты микроядер и хромосомных аберраций, что свидетельствовало о наличии «байстэндер» эффекта.
4.3.3.Как сравнить «байстэндер» эффект
иэффект прямого повреждения?
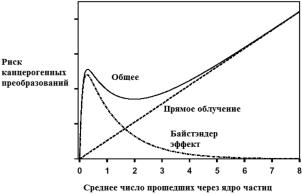
Данные, полученные при изучении РИБЭ, указывают на значительную разницу между «байстэндер» эффектом и эффектом прямого повреждения. Схематически на рис. 4.9 приведены для канцерогенных преобразований IN vitro вклады эффектов прямого повреждения, «байстэндер» эффекта и их сумма.
При достаточно высоких дозах эффект прямого повреждения убивает все клетки в облученной популяции.
В этом случае вклад РИБЭ незначителен и степень риска заболевания пря-
мо пропорциональна плотности облучения. С другой стороны, «байстэндер» эффект наблюдается при облучении малой части клеток, и повреждения обна-
руживаются среди тех клеток, которые даже не подвергались воздействию. Обычно от 1 до 30 % необлученной фракции клеточной популяции могут об-
наруживать повреждения, вызванные «байстэндер» эффектом. В этом случае РИБЭ преобладает над эффектом прямого повреждения.
При прямом повреждении процент погибающих клеток прямо пропорцио-
нален дозе, а при «байстэндер» эффекте наблюдается низкий порог дозы, за которым клеточный ответ представляет плато – дальнейшее повышение дозы не вызывает увеличения количества поврежденных клеток.
Существуют две основные теории, объясняющие наличие плато. Одна из них – есть уже существующая доля клеток в популяции, которая чувствитель- на к сигналам об облучении соседних клеток. А вторая – то, что вся популя- ция клеток отвечает на сигналы, но также одновременно генерирует подавля-
ющий сигнал, который ограничивает ответ определенной фракции клеток.
Рис. 4.9. Схема удельных вкладов прямого облучения и «байстэндер» эффекта в общее значе-
ние радиационного риска
181
Есть две основные области, где РИБЭ может играть важную роль. Первая связана с риском облучения низкими дозами естественного радиоактивного фона. Условная оценка облучения показывает, что естественный радиоактив-
ный фон является причиной 3 % всех случаев злокачественных опухолей, но является ли это преувеличением либо недооценкой – это частично зависит от предположений о биологическом действии на единичные клетки.
РИБЭ также может иметь значение при некоторых видах лучевой терапии. Например, при лечении рака точечное воздействие радиации не всегда может достигнуть цели, и тогда необлученные клетки опухоли поражаются с помо-
щью механизмов РИБЭ [364].
4.3.4. Исследования «байстэндер» эффекта in vivo
Радиационно-индуцированный «байстэндер» эффект не является исклю- чительно явлением IN vitro на клеточных культурах. Появляется все больше свидетельств того, что РИБЭ также может быть обнаружен IN vivo.
Так, было показано, что при накоплении α-источников в печени китайско- го хомячка все клетки ее обладают одинаковым риском индукции хромосом-
ных повреждений, даже если очень малая часть всей популяции подверглась облучению [365]. В другом исследовании при трансплантации отличающихся цитогенетическими маркерами облученных и необлученных клеток костного мозга в последующих клонах гемопоэтических стволовых клеток наблюда-
лась хромосомная нестабильность [366].
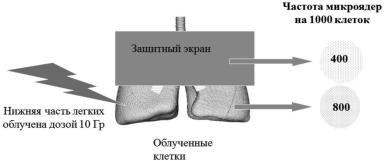
Врезультате облучения нижней части легкого крыс наблюдалась повы-
шенная частота микроядер в необлученной верхней части легких, которая снижалась при предварительном введении супероксид дисмутазы (рис. 4.10) [367]. Схожий эффект был описан у пациентов с хронической лейкемией [368]
ив костном мозге детей с хронической гранулоцитарной лейкемией в резуль-
тате облучения селезенки [356].
Вдругом исследовании форель подвергали облучению, а затем через не- сколько часов ее из аквариума пересаживали в аквариум к необлученной фо-
рели, а в этот аквариум пересаживали другую необлученную рыбу [369].
Рис. 4.10. Схема эксперимента по индукции РИБЭ IN vivo
182
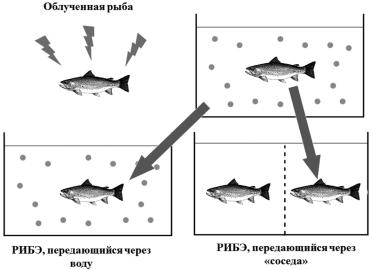
В результате у обеих интактных рыб наблюдалось повышение частоты ми-
кроядер по сравнению с контролем (рис. 4.11).
Особое значение РИБЭ имеет при фракционной терапии. Было показано, что культуральная среда от клеток, облученных несколькими фракциями, об-
ладала более выраженным цитотоксическим эффектом при переносе клеткамреципиентам по сравнению с средой от клеток, получивших одиночную рав- ную дозу облучения. Этот эффект противоречит наблюдаемому восстановле-
нию при прямом фракционном облучении культуры клеток – адаптивному ответу. Так, образование «байстэндер» факторов IN vivo может способствовать снижению «щадящего» эффекта, являющегося целью фракционной радиоте-
рапии [364].
В настоящее время в некоторых клиниках врачи борются с солидными опухолями путем пространственной фракционной радиотерапии, опосредо-
ванной формированием «байстэндер» эффекта, что способствует регрессии опухолей и снижению дозы обучения пациентов. Результаты этих и других исследований убедительно свидетельствуют, что РИБЭ существует не только исключительно в культурах, но и IN vivo.
При остром облучении выделение и эффект «байстэндер» факторов может быть больше и происходить в более короткие сроки. В то же время при хрони-
ческом низкодозовом облучении количество выделяемых в кровяное русло факторов может быть в очень малой концентрации, не способной вызвать кле- точное повреждение вне зоны локализации источника. Таким образом, на ос-
новании этих исследований можно сделать вывод, что «байстэндер» эффект, индуцированный малыми дозами, не будет обладать каким-либо эффектом вне облученной ткани и, следовательно, не будет иметь большое значение
IN vivo.
Рис. 4.11. Схема эксперимента по индукции РИБЭ IN vivo
183
Понимание эффектов радиации как скоординированного многоклеточного воздействия, поражающего не только облученные, но и необлученные клетки, позволяет определить вклад эффектов в необлученных клетках в оценку ра-
диационных рисков. В результате при оценке радиационного риска во всех моделях канцерогенеза однозначно должны приниматься во внимание не только мишенные, но и немишенные аспекты радиационного воздействия.
Очевидно, что будущие терапевтические методы будут включать восста- новление тканевой способности контролировать ответ на радиационное воз-
действие.
4.3.5. Механизмы радиоиндуцированного «байстэндер» эффекта
Повреждения в необлученных клетках могут быть объяснены возможно- стью взаимодействия клеток и передачей информации от одной клетки к дру- гой. Механизмы, используемые клетками для обмена информацией о радио-
активном поражении, до сих пор не ясны.
Есть свидетельства существования как минимум двух независимых путей передачи повреждения от облученной клетки необлученным: через межкле- точные взаимодействия и через клеточные факторы, секретируемые в культу-
ральную среду.
Всистемах, где есть прямой контакт клеток, по-видимому, важную роль играют межклеточные коммуникации и сигнал передается через щелевидные соединения (узкие каналы, имеющие приблизительно 2 нм в диаметре, кото-
рые связывают цитоплазмы двух смежных клеток и способствуют диффузии мелких молекул) [362].
Всистемах, где нет прямого контакта, взаимодействие происходит косвен-
ным путем, межклеточная передача информации может осуществляться (по крайней мере в некоторых случаях) химически активными радикалами кисло-
рода (ROS), вызванными действием ионизирующего излучения, или с помощью биологических веществ. Было показано, что культуральная среда от альфа-об-
лученных клеток способна индуцировать сестринские хроматидные обмены, увеличение генетических повреждений и снижение выживаемости клеток при
еепереносе необлученным клеткам [349]. Была выдвинута гипотеза, что об- лученные клетки выделяют цитотоксические факторы в культуральную сре-
ду, что способствует передаче сигнала необлученным клеткам.
Несмотря на то, что к настоящем моменту специфический фактор (сигнал) не был идентифицирован, возможный механизм может включать Ил-8, при-
нимающий участие в α-индуцированном «байстэндер» эффекте [357]. Другим возможным медиатором «байстэндер» эффекта является апоптоз
индуцирующий фактор (AIF), секретируемый митохондриями в ответ на окислительный стресс. Хотя AIF действует внутриклеточно, он может блоки- ровать выделение экстраклеточных цитотоксических факторов в культураль-
ную среду [370].
184
Имеются данные, что в процесс вовлекаются цитокины, которые выделя- ются клетками и играют специфическую роль в межклеточном взаимодей-
ствии [371].
Предполагается также, что повреждающий фактор обладает белковой при-
родой, так как он термолабилен, не теряет эффекта при замораживании, а при воздействии ингибиторами белков не формируется [351].
Показано также, что происходит подавление таких генов, как P53 и P21, во-
влеченных в процессы контроля клеточного цикла и индукции апоптоза [372]. Неизвестно, формируется ли в результате воздействия различных типов ионизирующей радиации один и тот же фактор, или сигналы различаются по
своей природе и эффекту.
4.3.6.Биологическая роль «байстэндер» эффекта
Внастоящее время дискутируется вопрос о биологической роли «байстэн-
дер» эффекта. Если данный эффект является повреждающим, увеличивающим число клеток, поврежденных одиночным радиационным треком, тогда не понятно, почему такой феномен сохранился в ходе эволюции? Зачем он нужен?
Предполагается, что главной функцией этого эффекта является уменьше-
ние риска трансформации клеток в многоклеточном организме, подвергшемся облучению. Можно предположить, что защитные системы организма не всег- да могут отреагировать на повреждения одиночных клеток внутри ткани, по- этому увеличение числа дефектных клеток необходимо для индукции и сти-
муляции систем, ответственных за уменьшение риска трансформации клеток
вмногоклеточном организме. При этом происходит репарация повреждений или апоптоз – удаление клеток, которые могли бы быть трансформированы, например, в раковые клетки.
Можно предположить также, что усиление повреждения («байстэндер» эффект) при воздействии малых доз необходимо для индукции адаптивного ответа.
Если РИБЭ осуществляет в организме защитные функции, становится по-
нятным, зачем эволюция его сохранила.
С другой стороны, нельзя исключить и того, что РИБЭ может усиливать эффект облучения, искажая зависимость доза–эффект.
При расчете радиационных рисков используются модели, основанные на прямом действии облучения на ядерную ДНК. Однако, если РИБЭ усиливает реакцию организма на воздействие ионизирующей радиации, то необходим перерасчет радиационных рисков, и в том числе риск канцерогенеза. При этом возможны различные формы нелинейности в зависимости доза–эффект
врайоне малых доз.
Знание этих закономерностей особенно важно для радиационной терапии рака, поскольку результат зависит от того, приводит ли РИБЭ к увеличению
185
числа трансформированных клеток или, наоборот, уменьшает их количество вследствие усиления летального эффекта.
Интерпретация «байстэндер» эффекта может быть различной с точки зре-
ния ожидаемого конечного результата. Если за параметр берется индукция трансформаций и/или мутаций, это может служить доказательством того, что опасность низких доз облучения выше, чем риск, вытекающий из зависимости доза–эффект.
Как следствие, с одной стороны, большое внимание должно уделяться оп-
тимизации процесса воздействия радиоактивного излучения в порядке предупреждения неоправданного облучения, которое может быть вреднее, чем предполагалось прежде.
С другой стороны, напротив, возрастающее количество апоптозов предпо- лагает возможный защитный эффект, принимая во внимание, что потенци-
ально пораженные клетки элиминируются из выжившей популяции.
4.3.7. Модификация «байстэндер» эффекта
Модификация РИБЭ химическими или физическими факторами (подавле- ние или усиление эффекта) еще больше усложняет зависимость биологиче-
ских последствий облучения от поглощенной дозы.
В частности, нами изучались эффекты некоторых радиопротекторных ве- ществ, обладающих высокой антирадикальной активностью (меланина, мела-
тонина и альфа-токоферола) на степень проявления «байстэндер» эффекта в клетках-реципиентах.
Выбор протекторов был обусловлен следующими данными. Так, меланин способен перехватывать и превращать в тепло все виды физической энергии – магнитную, электрическую, радиационную, звуковую, тепловую и т. д. Кроме того, меланин также является антиоксидантом, нейтрализует потенциально опасные свободные радикалы.
Существуют доказательства радиопротекторного действия меланина. Этот пигмент повсеместно распространен в живом мире. Он проявляет свое радио- протекторное действие на начальных стадиях облучения, предотвращая по-
вреждение ДНК и не влияя на репарационную систему.
Меланин очень эффективен при защите наследственных структур от му- тагенного действия малых доз радиации, при этом чем ниже доза радиацион-
ного воздействия, тем выше протекторное действие меланина. Так, меланин способен полностью предотвращать эффекты малых доз радиации порядка 0,2 Гр. Все это позволило предположить, что меланин может оказать воздей-
ствие на передачу сигналов от облученных клеток к необлученным [373]. Мелатонин – это нейрогормон шишковидной железы. Мелатонин секрети-
руется в основном ночью, свет подавляет его секрецию. В организме мелато- нин участвует в регуляции суточных и циркадных ритмов, температуры, сни-
жает эффект «реактивной» болезни и обладает противоопухолевой активностью.
186
Кроме того, мелатонин имеет антиоксидантные свойства и способен нейтра- лизовывать многие из известных радикалов, а также усиливать секрецию не-
которых противооксидантных ферментов.
Во многих исследованиях также было показано, что мелатонин обладает радиопротекторными свойствами.
Альфа-токоферол – витамин Е, известный своими радиозащитными свой-
ствами и антирадикальной активностью.
В качестве тест-системы использовалась культура клеток кератиноцитов человека, иммортализованных вирусом папилломы человека. Клетки-доноры РИБЭ облучали в малых дозах ионизирующей гамма-радиации. После выде-
ления «байстэндер» фактора в культуральную среду, ее фильтровали через 0,22 мкм фильтр и переносили необлученным клеткам-реципиентам, которые затем помещали в термостат и далее культивировали и обрабатывали в зави- симости от используемого метода анализа (микроядерный или колониеобра-
зующий тест) [373].
Для того чтобы вводимые в питательную следу протекторы не могли воз- действовать непосредственно на сами донорские клетки при облучении, за- щищая их, что могло бы способствовать уменьшению образования поврежда-
ющих факторов, радиопротекторы вводили через 1 ч после облучения, когда повреждающий фактор уже был сформирован, перед фильтрацией и перено-
сом среды необлученным клеткам-реципиентам.
Кроме того, необходимо было убедиться, что протекторы не проходят че-
рез фильтр и не попадают к клеткам – реципиентам, осуществляя обычный радиозащитный эффект. Для этого с помощью спектрофотометра сравнили оптическую плотность культуральной среды, среды с добавлением протекто- ров и среды, отфильтрованной от облученных клеток и протекторов. Интакт-
ная среда была идентична профильтрованной, что подтверждало отсутствие протекторов в переносимой клеткам-реципиентам среде.
Полученные данные показали, что перенос среды от облученных клеток необлученным способствует статистически значимому увеличению числа ми- кроядер и снижению выживаемости клеток-реципиентов по сравнению с кон- тролем, т. е. был выявлен РИБЭ. При введении радиопротекторов данный по-
вреждающий эффект был достоверно ниже (рис. 4.12, 4.13).
По результатам исследования, наибольшим защитным эффектом обладал мелатонин, наименьшим – альфа-токоферол [374]. Таким образом, было показано, что радиопротекторы способны уменьшать повреждающий эффект «байстэндер» факторов.
Таким образом, показана возможность уменьшения «байстэндер» эффекта с помощью радиопротекторов меланина, способного абсорбировать все виды физической энергии и обладающего высокой антирадикальной активностью, и мелатонина, также обладающего высокой антирадикальной активностью. Учитывая, что все использованные защитные вещества обладают выражен-
ными антирадикальными свойствами, можно предположить, что в механизме БЭ существенную роль играют факторы радикальной природы.
187
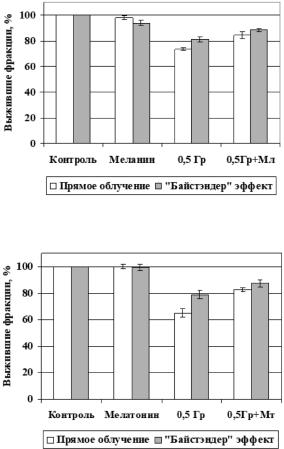
Рис. 4.12. Влияние меланина (Мл) на выживаемость клеток человека в результате прямого облучения и «байстэндер» эффекта
Рис. 4.13. Влияние мелатонина (Мт) на выживаемость клеток человека в результате прямого облучения и «байстэндер» эффекта
Поскольку предполагается, что главной функцией «байстэндер» эффекта является уменьшение риска канцерогенной трансформации клеток в организ- ме, подвергшемся облучению, полученные данные по уменьшению «байстэн- дер» эффекта с помощью меланина и мелатонина имеют большое практиче-
ское значение для радиационной терапии рака.
При расчете радиационных рисков используются модели, основанные на прямом действии облучения на ядерную ДНК. Однако, если «байстэндер» эффект усиливает реакцию организма на воздействие ионизирующей радиации, то необходим перерасчет радиационных рисков, и в том числе риск кан- церогенеза. Использование радиопротекторов, способных подавлять межкле-
точную передачу радиационных сигналов, позволит управлять этим явлением как в целях радиационной терапии рака, так и для оценки радиационных ри-
сков.
188
4.4. Современные проблемы биодозиметрии
Мутагенное действие ионизирующей радиации было доказано Дж. Мёлле-
ром в 1927 г. С тех пор прошло 90 лет, а самый главный вопрос о генетических последствиях облучения человека так и не решен. Для того чтобы определить частоту радиационно-индуцированных мутаций у дрозофилы, ученому при-
шлось синтезировать специальные тестовые линии, провести скрещивания облученных самцов с девственными самками и проанализировать второе или даже третье поколение. Что же касается человека, то его нельзя ни облучать, ни скрещивать, а исследовать частоту радиационных мутаций в потомстве людей, облученных по разным причинам, задача сложная.
Одним из подходов, позволяющих хоть как-то приблизиться к решению данного вопроса, является анализ цитогенетических повреждений, индуциру-
емых облучением в культивируемых клетках человека. При облучении in vitro одних и тех же образцов крови разными дозами ионизирующей радиации на-
блюдается строгая зависимость частоты аберраций от дозы (рис. 4.14, 4.15). Это дало основание для использования данного критерия в целях биодозиметрии.
Однако при обследовании населения Хиросимы и Нагасаки не было выяв- лено достоверных различий по уровню хромосомных перестроек между об-
лученными и не облученными группами людей. Результаты исследования in vivo оказались отличными от таковых in vitro.
После Чернобыльской аварии исследования по биодозиметрии получили широкое распространение, а сам термин «биодозиметрия» стал настолько по- пулярным, что часто использовался не по назначению. Так, в научной литера-
туре встречаются работы, в которых термином «биодозиметрия» называют определение количества радионуклидов в организме, а также суммарной или
Рис. 4.14. Зависимость доза–эффект для частоты маркерных аберраций (лимфоциты, 2–3-й митоз, облучение in vitro, по оси Х – доза, Гр; по оси Y – частота стабильных аберраций, %) [375]
189