

Радиобиология / Моссэ И. Б., Морозик П. М. Генетические эффекты ионизирующей радиации
.pdfВ течение 1998–2000 гг. исследовали генетическую радиочувствитель-
ность природных популяций дрозофилы из загрязненных радиацией районов Гомельской области (н. п. Ветка и Светиловичи; 24 Кu/км2) в сравнении с по-
пуляцией из Березинского заповедника (контроль). Выборки особей из этих популяций подвергали дополнительному однократному облучению в дозе 30 или 40 Гр. Резистентность особей к действию радиации проверяли по тестам рецессивных сцепленных с полом летальных мутаций (РСПЛМ), доминант- ных летальных мутаций (ДЛМ), включающих в себя эмбриональные леталь- ные мутации (ЭЛМ) и постэмбриональные летальные мутации (ПЭЛМ) со-
гласно стандартным методикам.
Как показали результаты исследований 1998 г., после острого облучения
вдозе 40 Гр частоты ЭЛМ, ПЭЛМ и ДЛМ в березинской популяции состави-
ли 63,1±0,9 %; 58,1±0,9 % и 84,5±0,7 %, соответственно, что было достоверно выше, чем в популяции из н. п. Ветка после острого облучения (ЭЛМ –
42,8±0,9 %; ПЭЛМ – 26,2±0,8 %; ДЛМ – 57,7±0,9 %). Частота индуцированных дополнительным облучением РСПЛМ в березинской популяции достигла 12,6±1,2 %, а в популяции из н. п. Ветка – 6,7±0,7 %. Эти данные свидетель- ствуют о большей устойчивости к дополнительному острому облучению по-
пуляций, отловленных в радиационно-загрязненных районах [217]. Результаты исследований, проведенных в 1999–2000 гг., оказались анало-
гичными предыдущим. В обоих случаях в качестве контроля использовали насекомых из Березинского заповедника. В 1999 г. дрозофил отлавливали
взагрязненном радионуклидами н. п. Светиловичи, а в 2000 г. – н. п. Ветка. При сравнении частот спонтанных мутаций выявлено, что в природных по-
пуляциях из загрязненных радионуклидами районов их уровень выше, чем
вконтроле. Однако при дополнительном остром облучении показатели ЭЛМ, ПЭЛМ и ДЛМ были выше в контрольной березинской популяции, чем в по- пуляциях из н. п. Светиловичи и Ветка после острого облучения. Эти резуль- таты позволяют сделать выводы о том, что уровень мутаций выше в популя- циях, постоянно подвергающихся хроническому воздействию малых доз ра-
диации, и при остром дополнительном облучении в высоких дозах радиации эти популяции являются более резистентными, т. е. у них формируется радио-
адаптация.
Для проверки специфичности адаптации использовали химический мута-
ген этилметансульфонат (ЭМС). Насекомых, отловленных из указанных выше районов, сравнивали по используемым генетическим параметрам до и после обработки ЭМС. Получено, что после обработки ЭМС в березинской популя- ции частоты ПЭЛМ и ДЛМ выросли в 1,5 раза, тогда как в популяции из за- грязненного радиацией н. п. Светиловичи, эти показатели остались без изме-
нения [217].
Аналогичная динамика выявлена также по показателю жизнеспособности. Под действием дополнительного облучения и этилметансульфоната жизне- способность особей в популяциях из радиационно-загрязненного н. п. Свети-
110
ловичи снижается меньше, чем в контрольной березинской популяции, что также свидетельствует о формировании адаптации к неблагоприятным эколо-
гическим факторам.
Таким образом, показано наличие адаптации природных популяций дро- зофилы из районов с повышенным радиационным фоном к мутагенному дей- ствию радиации. Выявлено, что природные популяции из загрязненных ради-
онуклидами районов стали более устойчивы к воздействию факторов не только физической, но и химической природы (ЭМС). Это свидетельствует о неспецифическом характере адаптации природных популяций дрозофилы из за-
грязненных радиацией районов Беларуси.
Адаптивная реакция популяции на радиационное воздействие не является простым механическим отбором более устойчивых генотипов. Этот процесс, сопровождающийся преобразованием генетической структуры популяций, приводит к формированию оптимальных генотипов, обладающих высокой устойчивостью к радиационному воздействию. Ген-энзимные системы явля-
ются наиболее удобными моделями при изучении влияния конкретных генов на приспособленность организмов.
В течение 1998–2000 гг. исследовали генетическую структуру природных популяций дрозофилы, обитающих в Березинском заповеднике и в радиаци- онно-загрязненных н. п. Ветка и Светиловичи, по соотношению частот гено-
типов и F- и S-аллелей локусов алкогольдегидрогеназы (Adh) и глицерофосфатдегидрогеназы (Gpdh), регулирующих баланс НАДН/НАД+, а также супе-
роксиддисмутазы (Sod), являющейся ключевым ферментом антиоксидантной защиты клеток и организма. Все популяции оказались полиморфными, в них были выявлены F-(быстрые) и S-(медленные) аллели изученных локусов. В популяциях, отловленных в н. п. Ветка и Светиловичи, частота гетерозигот FS
по локусам Adh и Gpdh, обладающих большей генетической пластичностью по сравнению с гомозиготами, была выше, чем в березинской популяции (по локусу Adh 0,30±0,05 по сравнению с 0,17±0,06 в контроле и по локусу Gpdh 0,50±0,05 по сравнению 0,38±0,05 в контроле).
Как известно, генетическая структура природных популяций находится
впрямой зависимости от экологических факторов, а гетерогенность популя- ций служит основой их адаптации к неблагоприятным условиям внешней сре- ды. Полученные результаты свидетельствуют о генетической перестройке по- пуляций дрозофилы под влиянием ряда экологических факторов, включая ра-
диационное воздействие.
Проанализирована также генетическая структура выборок из природных популяций, обитающих в районах с повышенным радиационным фоном, а также
вБерезинском заповеднике по локусу Sod. Наиболее ценным является обнаруженный факт вытеснения F-аллелей Sod и замена их на S-аллели в популяциях, подвергаемых в течение многих поколений хроническому облучению. S-форма этого фермента играет важную адаптивную роль и может обеспечить
радиорезистентность особей, поскольку известно, что супероксиддисмутаза
111
является эффективным радиопротектором на молекулярном, клеточном и ор-
ганизменном уровнях.
Выявлено, что частота S-аллеля этого локуса в популяциях из радиа-
ционно-загрязненных районов Ветка и Светиловичи (24 Ки/км2) составляет 0,60±0,05 и 0,64±0,06 соответственно, что существенно выше, чем в березин-
ской, где уровень S-аллеля 0,12±0,04. На основании собственных и литературных данных можно заключить, что S-аллозим Sod играет важную функциональную роль в адаптации популяций, обитающих в условиях повышенного радиационного фона.
Для оценки меры генетической изменчивости популяций была рассчитана гетерозиготность (Н) популяций по трем изученным локусам. В выборках из популяций н. п. Ветка и Светиловичи средняя гетерозиготность была значи-
тельно выше (0,47 и 0,43 соответственно), чем в березинской (0,25).
В популяциях из в н. п. Ельск, Наровля и Ветка, соотношения частот гено-
типов по изучаемым генам отличались от таковых в березинской популяции. Для всех ген-энзимных систем отмечено возрастание частоты гетерозигот, что обеспечивает популяциям большую пластичность в неблагоприятных экологических условиях обитания. Все популяции оказались полиморфными,
вних были выявлены F- (быстрые) и S- (медленные) аллели изученных локусов. В выборках из популяций, отловленных в н. п. Ветка и Светиловичи, ча- стота гетерозигот FS по локусам Adh и Gpdh была выше, чем в березинской
популяции [217].
Высокий уровень гетерозиготности дает адаптивные преимущества осо-
бям, но на популяционном уровне он может оказаться неблагоприятным из-за выщепления в поколениях большого количества неадаптивных генотипиче-
ских комбинаций.
Кроме того, в выборках из природных популяций, обитающих в радиа- ционно-загрязненных районах, выявлено возрастание частоты гетерозигот и S-аллельной формы супероксиддисмутазы, являющейся эффективным ра- диопротектором и обеспечивающей таким образом большую приспособлен- ность популяции к повышенному радиационному фону. Известно, что радио-
устойчивость SS-гомозигит локуса Sod превышает устойчивость особей с генотипами FF и FS. Таким образом, в экологически неблагоприятных условиях среды формируются генотипы с высокой приспособленностью и одновремен- но происходит элиминация менее приспособленных (неадаптивных) геноти-
пов, т. е. происходит обеднение генофондов популяций.
Следовательно, в условиях радиационного загрязнения преобразование ге-
нетической структуры популяций приводит к формированию оптимальных генотипов, обладающих более высокой устойчивостью к радиационному воз- действию и обеспечивающих тем самым выживание популяции в новых эко-
логических условиях.
Как известно, генетическая структура природных популяций находится
впрямой зависимости от экологических факторов, а гетерогенность популя-
112
ций служит основой их адаптации к условиям среды за счет постоянной эли- минации менее приспособленных генотипов и размножения генотипов, отно-
сительная приспособленность которых в данных условиях выше. Наиболее ценным является обнаруженный факт вытеснения F-аллелей Sod и замена их
на S-аллели в популяциях, подвергаемых в течение многих поколений хроническому облучению. S-форма этого фермента играет важную адаптивную
роль и может обеспечить радиорезистентность особей, поскольку известно, что супероксиддисмутаза представляет собой эффективный радиопротектор на молекулярном, клеточном и организменном уровнях.
Более резистентные особи способны быстро заполнять ареалы обитания, так что численность насекомых в радиационно-загрязненных районах не уменьшается. Более того, поскольку повышенная резистентность облучав-
шихся популяций является неспецифической, такие насекомые оказываются более устойчивыми к различным средствам химической борьбы с вредителя- ми. Эти факты позволяют прогнозировать возрастание численности насеко-
мых-вредителей сельского хозяйства в районах с повышенным радиационным фоном и рекомендовать использование для борьбы с вредителями сельского хозяйства другие методы (например, биологические и агротехнические).
Таким образом, изучение генетической радиочувствительности природ- ных популяций насекомых выявило изменения генетической структуры по- пуляций, обитающих в радиационно загрязненных районах, в сторону повы- шения их устойчивости к длительному воздействию облучения. Так, соотно-
шение аллелей аллозимных локусов в популяциях дрозофилы сдвигается в пользу генотипов, наилучшим образом приспособленных к повышенному радиационному фону. При дополнительном остром облучении в высоких до-
зах генетический ответ популяций дрозофилы из радиационно загрязненных районов оказывается меньшим, чем из контрольных, что свидетельствует о большей генетической радиоустойчивости этих популяций. Выявлен не- специфический характер такой резистентности – популяции из районов с по-
вышенным радиационным фоном оказываются более адаптированными и к воздействию химического мутагена ЭМС. Адаптация популяций сопровождается гибелью чувствительных генотипов и обеднением генофонда.
Исследования природных и экспериментальных популяций разных видов, подвергающихся длительному воздействию ионизирующей радиацией, пока- зали, что в условиях повышенного радиационного фона в популяциях возрас-
тает частота морфологических нарушений и мутаций различных типов. В то же время усиливаются процессы меланизации, способствующей повышению устойчивости особей, и происходит постепенный отбор более резистентных генотипов, обеспечивающих адаптацию популяций к повреждающему дей- ствию облучения. Происходит элиминация менее приспособленных геноти-
пов и образование комбинаций генотипов, относительная приспособленность которых в данных условиях существования выше.
113
Таким образом, увеличение радиоустойчивости популяций при хрониче-
ском или периодическом облучении в определенных условиях возможно для различных видов живых существ.
Можно сделать вывод, что в природных популяциях, а также и в экспери- ментальных при жестком естественном отборе и при интенсивном воздей-
ствии селектирующего агента адаптация облучаемых популяций происходит довольно быстро. В лаборатории можно создать такие условия облучения и действия естественного отбора, при которых будет наблюдаться длительное равновесие между давлением мутаций и отбором. При этом длительность пе-
риода формирования адаптации популяций зависит от условий эксперимента. Понятно, что чем выше плодовитость и чем быстрее смена поколений у данного вида, тем легче может он приспособиться к воздействию ионизиру-
ющей радиации даже в высоких дозах. Быстрое размножение выживших осо-
бей позволяет заполнить ареал обитания популяцией, резистентной к данной дозе облучения.
Возникновение сверхрезистентных популяций особенно характерно для простейших, и такие примеры хорошо известны для вирусов и бактерий. На- пример, в воде атомного котла реактора в Лос-Аламосе живут и размножают-
ся бактерии Мicrococcus radiodurans при дозах 104–105 Гр.
Популяции насекомых также могут восстановить свою численность, даже если в результате неблагоприятных условий существования их численность резко сокращается (т. е. популяция проходит сквозь «бутылочное горлышко»). Например, популяции многих видов, обитающие на океанических островах, хотя и насчитывают в настоящее время миллионы особей, происходят от од-
ной или нескольких особей, когда-то давно случайно попавших туда. Некоторые популяции диких животных также могут восстанавливаться
после гибели подавляющей части особей (в частности, такие примеры извест- ны для популяций грызунов). Однако, чем ниже плодовитость и больше дли-
тельность поколений, тем труднее восстанавливается численность популяции. Совершенно особая ситуация складывается для популяций человека, где естественный отбор потерял ведущее значение и где число поколений в пре-
делах больших исторических отрезков времени очень невелико. Абсолютно ясно, что для человека такая адаптация, связанная с гибелью большой части популяции, невозможна и недопустима.
2.4.2. Распределение особей в популяции по признаку радиочувствительности
Радиочувствительность – количественный признак, такой как рост, вес, плодовитость и т. д. Одной из особенностей количественных признаков явля-
ется то, что кривая распределения особей с разной радиочувствительностью в популяции представляет собой колоколообразную форму. Для большинства
114
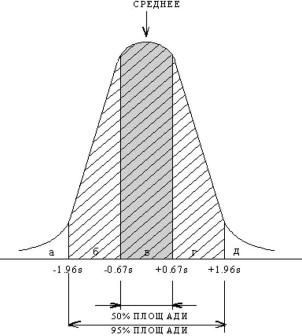
особей характерны промежуточные значения данного признака, и лишь у не- большой части особей обнаруживаются крайние его значения (рис. 2.13). Ма- тематическая кривая отражает нормальное (или биномиальное) распределе- ние. При этом 50 % популяции (или результатов наблюдений) попадает в ин-
тервал, заключенный между значениями –0,67s и + 0,67s, т. е. в зону среднего значения признака: х±0,67s (более темный участок графика), 95 % выборки (или популяции) находится в интервале х±1,96s (заштрихованный участок графика), и только 5 % популяции оказываются в пределах от х±1,96s до х±3s и разделяются на сверхчувствительную фракцию (2,5 %) и сверхрезистент-
ную (2,5 %). Это значит, что чем значение признака дальше от среднего, чем для меньшей доли популяции он характерен.
Таким образом, число организмов или клеток, чувствительных к повреж-
дающему агенту, возрастает не просто пропорционально дозе воздействия, а согласно кривой нормального распределения, в результате чего наблюдается резкое увеличение числа чувствительных генотипов на границе а и б и такой же резкий спад на границе г и д (рис. 2.13).
Наличие сверхчувствительной фракции в популяциях клеток и организ-
мов объясняет противоречивые мнения относительно порога биологического действия ионизирующей радиации. Действительно, если исследуются неболь- шие выборки из популяций или используются не очень чувствительные те- сты, то кажущийся порог в действии облучения выявляется на рубеже интер-
Рис. 2.13. Нормальное распределение
115
валов б и в. Чем дальше влево от этой границы, тем меньше количество осо- бей (или клеток), чувствительных к изучаемой дозе, поэтому чем чувстви-
тельней тесты и чем больше анализируемая выборка, тем при более низких дозах удается обнаружить биологические (в частности генетические) эффекты, что свидетельствует об отсутствии порога в генетическом действии радиации.
Очевидно, что малочисленная сверхчувствительная фракция в популяци- ях страдает раньше всех других от загрязнения окружающей среды радиону- клидами, так же как и другими повреждающими агентами. Что касается по- пуляций человека, то очевидно, это и есть та дань, которую человечество пла-
тит за цивилизацию.
Популяции разных видов живых организмов могут адаптироваться к вред-
ным воздействиям в высоких дозах, таких как в, при которых гибнет 75 % особей, а некоторые популяции способны восстановить свою численность даже после воздействия в дозе г, губительной для 97,5 % особей. Такая адапта-
ция достигается ценой гибели большей части популяции за счет размножения выжившей самой резистентной фракции.
Часто эпидемиологические исследования обнаруживают неравномерное распределение заболеваний по территориям. Например, в одной деревне вы- являются сразу несколько лейкозов. Это не всегда означает, что жители дан- ной деревни подверглись облучению в более высокой дозе. Этот феномен мо-
жет быть обусловлен более высокой радиочувствительностью заболевших, вероятно имеющих общие генетические корни.
При исследовании ликвидаторов иногда обнаруживается, что одни и те же признаки лучевой болезни появляются у лиц, получивших дозы ионизирующей радиации, различающиеся весьма существенно. Из этого факта некото- рые ученые делают вывод о неправильном вычислении дозовых нагрузок, за-
бывая о генетических различиях индивидуумов.
Для выявления риска малых доз радиации были обследованы популяции людей, проживающих в районах с повышенным радиационным фоном. На- пример, в штате Керала в Индии уровень естественной радиоактивности ра- вен 4–5 мЗв в год, а в штате Айова и Иллинойс в США концентрация ра-
дия-226 в питьевой воде составляет в среднем 0,17 Бк на литр. Достоверных отклонений в состоянии здоровья людей, проживающих на этих территориях, не выявлено [218]. Однако можно предположить, что сверхчувствительная фракция популяций, первоначально поселившихся в таких местах, была эли- минирована в ходе естественного отбора, а оставшаяся часть дала начало но-
вой, более резистентной популяции. Если же в эти места будут переселены люди из других популяций, то влияние высокого радиационного фона может привести к значимым для переселенцев биологическим последствиям.
Учитывая биномиальное распределение по признаку радиочувствительно- сти особей внутри популяции, можно разработать новые подходы к определе-
нию предельно допустимых уровней воздействия радиации.
116
Согласно такому распределению большая часть популяции обладает сред-
ним значением радиочувствительности. Чем более радиочувствительны или радиорезистентны особи, тем реже они встречаются в популяции (рис. 2.13). Чем более правое положение на рисунке занимают особи, т. е. чем они более устойчивы к радиации, тем большую дозу могут они перенести без вредных последствий. Поэтому безвредные для разных фракций популяции дозы ради-
ации на этом графике будут возрастать от а к д.
Конечно, на современном этапе невозможно избавить живую природу от воздействия неблагоприятных факторов в таких малых дозах, как а. При этом повреждается лишь 2,5 % популяции. Что же касается дозы б, то она губи-
тельна уже для 25 % особей. Этот резкий переход от а к б, очевидно, и должен быть пределом допустимых доз облучения, во всяком случае, для популяций человека.
Учет популяционно-генетических аспектов особенно важен для вычисле- ния индивидуального радиационного риска. Например, при лечении злокаче-
ственных заболеваний, таких как раковые опухоли, и особенно при лейкозах, необходимо очень точно определить дозы радиационно-терапевтического воз- действия. Эффективные дозы для разных лиц могут различаться очень силь-
но, а ошибки могут привести к летальному исходу.
2.4.3. Механизмы адаптации популяций
Одним из возможных путей формирования адаптации популяций к хро- ническому воздействию повреждающих факторов является отбор более рези- стентных генотипов. Известно, что популяции полиморфны по генам, контро- лирующим радиочувствительность. И если в обычных условиях особи, несу- щие гены повышенной радиоустойчивости, не имеют селективного преиму-
щества, то при хроническом облучении такие генотипы приобретают особую ценность и закрепляются отбором.
Каковы могут быть причины повышенной резистентности того или иного генотипа? Устойчивость к облучению может быть обусловлена увеличением содержания в клетках естественных радиопротекторов. Предполагается, что в условиях облучения происходит отбор мутантов, синтезирующих тиолы, больше, чем в контрольной популяции [211]. Для проверки этой гипотезы было определено содержание эндогенных тиолов у ряда природных радиоре- зистентных и чувствительных штаммов хлореллы и обнаружена четкая кор- реляция между радиочувствительностью и концентрациями внутриклеточ-
ных тиолов [211].
При воздействии мутагенов, и в частности облучения, в растениях проис-
ходит возрастание количества витаминов. Экспериментально были получены данные о существовании корреляции между эндогенным содержанием ауто-
антимутагенов и устойчивостью к действию естественных и искусственных мутагенов [212].
117
Причиной повышенной устойчивости организма может быть более эффек- тивное функционирование систем репарации. Так, при исследовании устойчи- вости популяций хлореллы к хроническому воздействию химического мута-
гена этиленимина было установлено, что резистентные штаммы хлореллы характеризуются более высокой эффективностью систем репарации, чем чув-
ствительные [219].
Адаптация популяций может осуществляться не только за счет отбора су- ществовавших в ней резистентных генотипов, но и за счет формирования ге- нотипа, наилучшим образом приспособленного к новым экологическим усло- виям. В работе [220] при биохимическом изучении соотношения специфиче- ских локусов в генофонде популяций пресноводного моллюска было выявле-
но, что повышенная устойчивость популяций возникает не из-за появления и размножения новых мутантных форм, а за счет формирования оптимально- го генотипа. В то же время под действием мутагенных факторов в популяци- ях могут возникать новые мутации повышенной резистентности. Такие мута- ции будут закрепляться в ходе естественного отбора как имеющие преимуще-
ство в новых условиях.
Разновидностью естественного отбора в популяциях является половой от-
бор. Действие его изучено в основном на популяциях насекомых. Например, вытеснение из популяций дрозофилы мутации vestigial, фенотипически про- являющейся в недоразвитии крыльев, осуществляется не только за счет низ- кой конкурентной способности мутантных особей по сравнению с нормаль- ными (например, конкуренция за пищу и др.). Помимо этого, существует по- веденческая компонента, заключающаяся в ассортативном спаривании в поль- зу особей «дикого» типа, проявление которого связывается со специфически-
ми феромонами, выделяемыми нормальными особями. При этом крылатые самцы безразличны к фенотипу самок, но крылатые самки предпочитают крылатых самцов [221].
Аналогичным образом вытесняется из популяции и мутация white (белые глаза). Низкая половая активность белоглазых самцов приводит к тому, что при исходном соотношении генов 1:1 полная элиминация мутации white про-
исходит к седьмому поколению [222] (столько же поколений требуется и для вытеснения мутации vestigial [221]). В то же время разные пигментные мута- ции по-разному влияют на конкурентную способность их носителей. Так, бе- лый цвет глаз снижает приспособленность особей, а желтый цвет тела (мута-
ция yellow) не влияет на конкурентные взаимоотношения [223].
У бабочек рода Golias самки осуществляют выбор партнера по «летатель-
ной активности». Особи, имеющие генотипы, обусловливающие способность к продолжительному полету, чаще других попадают в число партнеров по скрещиванию. Более того, при повторном скрещивании самки выбирают сам-
цов с «лучшим» генотипом чаще, чем при первом спаривании [224].
Таким образом, половой отбор может благоприятствовать генам, обеспе-
чивающим общее адаптивное преимущество. Материалом для естественного
118
отбора являются также мутации, возникающие при изменении окружающей среды.
Вгеографически изолированных популяциях дрозофилы наблюдаются одновременные изменения сайтспецифичной мутабильности, которые объяс- няются флуктуациями частот генов-мутаторов. Предполагается, что при гло-
бальных изменениях условий среды, таких как пандемия или введение новых мутагенов, селективная выживаемость устойчивых мутантов создает преиму- щество для генов-мутаторов, вызывающих мутации соответствующих локу-
сов [225].
Вряде исследований проводилось подробное выяснение причин радиоустойчивости популяции дрозофилы, долгое время подвергавшейся облуче- нию [226, 227]. Вначале было установлено, что повышенная радиорезистент-
ность этой популяции обусловлена устойчивостью стадии 7 ооцитов. Затем
спомощью дальнейшей селекции получена еще более радиоустойчивая попу- ляция и изучена генетическая система, ответственная за ее повышенную ра- диорезистентность. Показано, что радиационно устойчивая мутация 1, лока-
лизованная в Х-хромосоме, контролирует процесс образования доминантных и рецессивных летальных мутаций (ДЛМ и РЛМ соответственно). Мутация 2, расположенная в аутосоме 2, независимо от мутации 1 снижает вероятность образования ДЛМ и РЛМ и частоту потерь Х-хромосом, она снижает также вероятность межхромосомных обменов и нерасхождения хромосом. Мутация 3 (аутосома 3) независимо от двух предыдущих контролирует радиорезистент- ность в отношении ДЛМ, РЛМ и интеробменов с нерасхождениями. Установ- лено, что мутация 3 связана с одиночным рецессивным геном, локализован-
ным в 3-й хромосоме в положении 49.8.
Дальнейшие исследования радиорезистентности двух изучаемых популя-
ций показали, что она наследуется полудоминантно, а радиочувствительность гибридов между этими популяциями соответствует радиочувствительности родительских популяций [228].
Установлено, что адаптивная эволюция на основе рецессивных мутаций невозможна и благоприятная мутация, вызывающая увеличение резистентно-
сти популяции, должна быть доминантной или хотя бы полудоминантной [229]. Огромные преимущества в условиях длительного воздействия поврежда-
ющего фактора дает особям амплификация генов, ответственных за устойчи-
вость к этому фактору. Так, после многошаговой селекции с возрастающими концентрациями боррелидина были получены линии клеток яичника китай-
ского хомячка с резистентностью к боррелидину в 1000 раз большей, чем у родительской линии [230]. В работе показано, что эта устойчивость обуслов- лена амплификацией гена, кодирующего треонил-тРНК синтетазу, ингибито-
ром которой является боррелидин.
Врезультате цитогенетических исследований было выявлено, что линия клеток мышиной меланомы обладает устойчивостью к адриамицину, в 50 раз
119