

8.5. Моделирование на животных
Генетические исследования животных, в отличие от человека, обладают более широким спектром возможностей (скрещивания, селекция, манипуляции со средой, возможность вмешательства в функционирование организма для изучения внутренних механизмов и т.п.).
Благодаря значительной общности в организации геномов млекопитающих и человека, а также сопоставимости функционирования их нервной системы, в психогенетике используются методы моделирования на животных различных психических состояний, заболеваний, элементарных форм научения и др.
Модельные эксперименты на животных широко используются при изучении наследственности алкоголизма, эпилепсии, болезни Альцгеймера, агрессивности, склонности к беспокойству и страху, способности к научению.
Словарь терминов
Геномика
Структурная геномика
Функциональная геномика
Протеомика
Полиморфизм
Однонуклеотидный полиморфизм
Хромосомный полиморфизм
Генетический маркер
Сцепление
Кроссинговер
Рекомбинация
Анализ сцепления
Группа сцепления
Плейотропный эффект действия гена
Генетическая карта
Сантиморганида
Мера "генетического расстояния"
Локусы количественных признаков (ЛКП)
Главные гены
Неравновесное сцепление
Анализ ассоциаций
Ген-кандидат
Генетическая инженерия
Рекомбинантные ДНК
Обратная транскрипция
Полимеразная цепная реакция (ПЦР)
Геномная ДНК
Рестрикция
Секвенирование
Блот-гибридизация
Гель-электрофорез
Альтернативный сплайсинг
Экзон
Интрон
Трансгенные животные
Мыши-нокауты
Вопросы для самопроверки
Что такое геномика?
Какие направления геномики вы знаете?
Что вы знаете о программе "Геном человека"?
Что такое геномные полиморфизмы?
Что такое однонуклеотидные полиморфизмы?
Какие гены называются сцепленными?
Каков основной принцип анализа сцепления?
Что такое генетические карты?
Как измеряется расстояние между генами?
Что такое картирование генов?
Какие генетические маркеры могут использоваться при анализе сцепления?
Для каких признаков анализ сцепления имеет смысл?
Что такое локусы количественных признаков (ЛКП)?
Какие методы картирования ЛКП вы знаете?
Что такое главные гены?
Что такое метод ассоциаций в генетике?
Что такое ген-кандидат?
Дайте общее представление о современном направлении в генетике, называемом генной инженерией, и подумайте о его возможностях для изучения генетики поведения.
Какие методы прямого анализа ДНК используются в генетике поведения?
Что такое секвенирование ДНК?
Что такое методика генного нокаута и каковы ее перспективы в изучении поведения?
Что такое плейотропный эффект?
Почему в психогенетике приходится прибегать к моделированию на животных?
Почему, изучая генетику поведения животных, мы можем косвенно судить о генетике поведения человека?
Какие конкретные примеры моделирования на животных вы можете привести?
Что вы знаете о влиянии раннего опыта на обучаемость животных?
Какие аналогии между генетикой обучаемости у животных и генетикой интеллекта человека можно провести?
Почему генетически обусловленные различия между "умными" и "глупыми" животными проявляются не во всех условиях среды?
Можно ли в экспериментах с животными идеально уравнять условия среды?
Раздел V. Психогенетика и развитие тема 9. Генотип и среда в индивидуальном развитии
9.1. Концепция нормы реакции и развитие
9.2. Как среда может взаимодействовать с генотипом в процессе развития?
9.3. Молекулярные аспекты развития
9.4. Морфогенез нервной системы и факторы, влияющие на этот процесс
9.5. Роль эмбрионального и неонатального опыта в развитии поведения
9.6. Родительские эффекты в развитии
9.7. Принцип системности и диалектика взаимодействия генотипа и среды в развитии
Генетика поведения человека лишена возможности экспериментировать с объектом своего исследования и вынуждена пользоваться естественными "экспериментальными" ситуациями, имеющимися в природе и социуме (близнецы, приемные дети и т.п.), поэтому и изучение развития здесь ведется на популяционном уровне. Основная часть выводов генетики поведения носит статистический, популяционный характер, поскольку касается не реального взаимодействия генотипа и среды в процессе развития конкретного фенотипа, а имеет отношение к компонентам дисперсии фенотипов в популяции. Поэтому, говоря о природе генотип-средовых взаимовлияний в индивидуальном развитии, мы будем опираться не столько на данные генетики поведения человека, сколько на косвенные данные смежных дисциплин: генетики поведения животных, молекулярной генетики, эмбриологии, нейробиологии развития, физиологии и т.д.
9.1. Концепция нормы реакции и развитие
Популяционный подход к оценке наследуемости особенностей поведения не позволяет описывать процессы взаимодействия генотипа и среды в индивидуальном развитии. Когда в результате психогенетических исследований, проведенных, скажем, на близнецах или на приемных детях, признак относят к наследуемому, это не значит, что он наследственно детерминирован в общепринятом смысле этого слова. На первый взгляд, звучит парадоксально. Однако давайте разберемся. Мы уже много раз подчеркивали, что психогенетические исследования ведутся в основном на популяционном уровне. Когда на основании коррелирующего поведения у родственников популяционные генетики делают вывод о наследуемости признака, это не означает, что индивидуальное развитие данного поведения обусловлено исключительно генетическими причинами. Высокая наследуемость свидетельствует лишь о том, что разнообразие индивидов в популяции в значительной степени связано с генотипическими различиями между ними. Имеется в виду, что процент индивидов, обладающих данным признаком в популяции потомков, может быть предсказан, исходя из знаний о родительской популяции. Однако значение показателя наследуемости ничего не говорит о последовательности событий в индивидуальном развитии признака и о том, какой конечный фенотип будет результатом развития конкретного индивида. В этом смысле признак с высокой оценкой наследуемости не является детерминированным генотипом, хотя такие интерпретации часто встречаются даже в публикациях специалистов. Это совсем разные вещи - разделить источники вариативности в популяции на генетические и средовые или искать генетические и средовые причины, лежащие в основе онтогенетического формирования конкретных фенотипов. Вспомним наш вымышленный пример с приемными детьми, которые попали на воспитание в семьи с более высоким социоэкономическим уровнем и оказались по своему абсолютному уровню умственного развития ближе к приемным родителям. Изобразим эту ситуацию в графической форме, допустив существование линейной зависимости между параметрами среды и уровнем интеллекта (на самом деле зависимость скорее логарифмическая, (рис. 9.1).
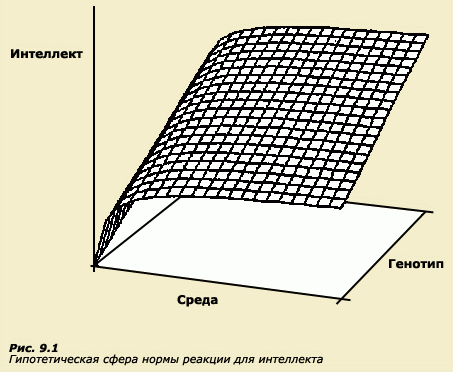

Мы получим серию индивидуальных графиков "нормы реакции" для 5 генотипов (рис. 9.2). Обозначим генотипы каждого из пяти детей цифрами 1, 2, 3, 4, 5 (ранговые места в порядке возрастания фенотипической выраженности признака) и рассмотрим их фенотипы в двух диапазонах среды - Е1 (обедненная) и Е2 (обогащенная). В нашем идеальном примере все дети, перейдя из обедненной среды в обогащенную, дали определенный прирост интеллекта, то есть фенотип каждого ребенка улучшился за счет чисто средовых причин. Однако ранговые места детей сохранились в соответствии с их генотипами. Ребенок, который в обедненной среде демонстрировал самый высокий интеллект, и в обогащенной оказался выше всех по результатам, и наоборот. Эта схема демонстрирует как бы соотношение между данными популяционно-генетических исследований и индивидуальным развитием. В данном примере высокая наследуемость признака проявляется в сохранении ранговых мест детей, но вместе с тем признак не является жестко детерминированным генотипом в том смысле, что конечный фенотип может быть изменен под влиянием изменений в условиях среды. Таким образом, даже при 100%-ной наследуемости, как она понимается в генетике поведения, есть возможность для влияния среды на формирование признака в индивидуальном развитии. Такой подход и соответствует генетическим представлениям о норме реакции. Вспомним, что наследуется не признак, а именно норма реакции. О норме реакции в данном разделе следует поговорить особо. Во многих учебниках генетики, в школьном курсе биологии и других книгах часто под нормой реакции понимают пределы, которые генотип кладет формированию фенотипа. Такое понимание нормы реакции, на наш взгляд, менее продуктивно, чем то, которого придерживаемся мы в ходе изложения материала. Вспомним, что мы определили норму реакции как специфический характер реакции генотипа на изменения среды. Введение в определение нормы реакции понятия предела вполне объяснимо, поскольку в обычных стандартных условиях развития, действительно, генотипы ограничивают возможности развития фенотипов. Например, люди с хорошими генетическими задатками для развития интеллекта при прочих равных всегда будут опережать людей с плохими задатками. Считается, что среда может сдвигать конечный результат развития, но в пределах диапазона, который генетически детерминирован. Но, в действительности, это ложная посылка, поскольку мы никогда не можем быть уверены, что признак достиг максимального развития, возможного для данного генотипа. Характер фенотипических проявлений генотипа не может быть протестирован для всех возможных сред, поскольку они неопределенно вариабельны (Dobzhansky Т., 1995). В отношении человека мы не имеем возможности не только произвольно контролировать параметры среды, в которой происходит развитие, но часто, анализируя средовые влияния на признак, затрудняемся даже в выборе тех параметров, сведения о которых необходимо получить, особенно если речь идет о поведенческих характеристиках. Современная психобиология развития поставляет все больше данных о значительных возможностях среды, в частости раннего опыта, в том числе эмбрионального, влиять на активность генов и структурное и функциональное формирование нервной системы. Таким образом, если в традиционной среде создается иллюзия того, что существуют пределы для формирования фенотипа, то мы не можем быть уверены, что развитие, в процессе которого генотип будет подвергаться необычным, нетрадиционным воздействиям, не приведет к возникновению таких особенностей поведения, которые в обычных условиях при данном генотипе были бы невозможны. Таким образом, более правильно считать, что пределы фенотипа непознаваемы. Многие с интересом следят за публикациями о нетрадиционных методах воспитания младенцев, а некоторые родители испытывают их на своих детях. Кто-то пытается вырастить музыканта, начиная с внутриутробного периода, когда мать, вынашивающая ребенка, с помощью нехитрых приспособлений обеспечивает своему плоду прослушивание музыкальных произведений или сама поет колыбельные еще не родившемуся ребенку. Кто-то рожает в воде и затем плавает с новорожденным в ванне или бассейне. Кто-то увлекается динамической гимнастикой и закаливанием. Все чаще в роддомах младенца в первые минуты жизни не отлучают от матери, как это традиционно делалось раньше, а еще до перерезания пуповины кладут к ней на живот, обеспечивая столь естественный контакт матери и новорожденного. Все эти "эксперименты" есть не что иное, как воздействие нетрадиционного (для данного периода развития общества) раннего опыта на плод и новорожденного, и эти воздействия не лишены смысла, поскольку интенсивно формирующаяся нервная система, от которой, в конечном счете, и будет зависеть наше поведение и все высшие психические функции, очень податлива к воздействиям именно в ранний период онтогенеза. Что же известно на сегодняшний день о влиянии раннего опыта, то есть среды, на развитие нервной системы и может ли эта среда влиять непосредственно на работу генетического аппарата? Иными словами, это вопрос о том, какими знаниями о процессе взаимодействия генотипа и среды в индивидуальном развитии мы располагаем.
9.2. Как среда может взаимодействовать с генотипом в процессе развития?
Понятно, что результат развития - фенотип - зависит от совместного действия генов и среды. Гены и признаки связаны сложной сетью путей развития. Все индивидуальные различия, которыми занимаются дифференциальные психологи и психогенетики, являются результатом обстоятельств развития конкретных индивидов в конкретных средах. Часто индивиды, воспитанные в явно различающихся средах, имеют много общего. И наоборот, сиблинги, воспитывающиеся в одной семье, казалось бы при сходных обстоятельствах, за счет тонких различий в условиях воспитания и развития реально будут испытывать весьма различные воздействия как физической, так и социальной среды. Это справедливо даже для генетически идентичных МЗ близнецов. Таким образом, процесс взаимодействия со средой сложен и неоднозначен. Отметим также, что психологи и другие исследователи часто пользуются термином "взаимодействие" в статистическом смысле, когда исследуется взаимодействие отдельных факторов в продуцировании какого-либо измеряемого эффекта. Подчеркнем, что статистическое взаимодействие факторов и взаимодействие генов и среды в индивидуальном развитии - это совершенно разные вещи. Их не следует путать. (Более подробно о статистическом взаимодействии факторов наследственности и среды см. в темах 4 и 6.) Для нас вполне привычной является формулировка, в которой утверждается, что проявление фенотипа является результатом взаимодействия генотипа со средой в процессе развития. Однако если вдуматься в это утверждение, оно не покажется столь очевидным. Ведь взаимодействие предполагает, что его участники вступают в контакт, соприкасаются. На самом деле наш генотип, то есть генетический аппарат, спрятан глубоко внутри клетки и отделен от внешней среды не только покровами тела, но и клеточной и ядерной оболочками. Как же внешняя среда может взаимодействовать с генетическими структурами? Ясно, что гены и окружающий мир непосредственно не соприкасаются. С внешней средой взаимодействует организм в целом; гены же взаимодействуют с различными биохимическими субстанциями внутри клетки. А вот различные клеточные субстанции могут испытывать влияние внешнего мира. Рассмотрим, что известно об этих процессах сегодняшней науке. Для этого снова придется обратиться к молекулярной генетике и более подробно рассмотреть, как функционируют гены, поскольку в предыдущем изложении мы лишь констатировали, что главной функцией гена является кодирование информации, необходимой для синтеза специфического белка.
9.3. Молекулярные аспекты развития
9.3.1. Фенотип на клеточном уровне
9.3.2. Транскрипция, трансляция и факторы, влияющие на эти процессы
9.3.3. Ранние гены и их роль в развитии
9.3.4. Гормоны и их роль в генетической регуляции
9.3.5. Регуляторная роль G-белков
9.3.1. Фенотип на клеточном уровне
В настоящее время подчеркивается, что необходимо разделять роль ДНК в передаче наследственной информации и в функционировании клетки (Michel G.F., Moore C.L., 1995). Первая обеспечивается способностью ДНК к точной репликации и существованием клеточных механизмов для попадания этих репликаций в гаметы. Вторая имеет непосредственное отношение к тому, что изучается как взаимоотношение между геном и признаком. По мере того как быстроразвивающаяся молекулярная генетика интенсивно накапливает данные, взгляд на ДНК как на некий код для хранения информации о признаке все более меняется. Когда исследуются детали функционирования клетки, оказывается, что существует множество промежуточных ступеней между ДНК и признаком, и множество агентов являются участниками превращения генетической информации в признак. ДНК является лишь одним из многих звеньев в динамической иерархически организованной системе. Для психолога понятие "фенотип" обычно ассоциируется с понятием анатомического или поведенческого признака. Среда для психолога включает различные факторы вне организма. Это и физическая среда (различные виды энергии, материальные объекты), и социальная среда (социальное окружение). Для генетика же среда есть все внешнее по отношению к генам, или ДНК. В этом смысле среда в первую очередь включает различные химические субстанции, продуцируемые клеткой или ее окружением, а также продукты метаболизма переваренной пищи. В свою очередь, понятие "фенотип" для молекулярной генетики ассоциируется с фенотипом клетки, а именно, ее белками. Белки являются наиболее важными физико-химическими субстанциями внутри клетки, обеспечивая ее структурные элементы и выполняя роль катализаторов и регуляторов всех аспектов жизнедеятельности клетки. Белки являются крупными молекулами, составленными из скрученных волокон, называемых полипептидами. Полипептиды составлены, в свою очередь, из рядов различных аминокислот. Типы, количество и последовательность аминокислот в белках определяют их физико-химическое строение и другие особенности. Эти особенности определяют функциональную роль белка (структурный белок, фермент, регулятор и т.д.). Генетический код, о котором мы говорили при изложении основ общей генетики, обеспечивает четкую параллель между последовательностью нуклеотидов в ДНК и последовательностью аминокислот в полипептиде. Мы уже знаем, что кодон - это три нуклеотида (триплет), кодирующие одну аминокислоту. Одна и та же аминокислота может кодироваться более чем одним кодоном. Например, кодон ГГЦ (гуанин, гуанин, цитозин) кодирует аминокислоту глицин. Эта же аминокислота кодируется еще тремя кодонами. Четыре нуклеотида, взятые по три, дают 64 сочетания. Большинство кодонов (61 из 64) кодируют аминокислоты. Оставшиеся три используются, чтобы отмечать конец кодируемой последовательности в гене. Генетический код был сначала исследован на прокариотах (доядерных организмах), в частности, бактерии Escherichia coli. У этих организмов порядок кодонов в молекуле ДНК точно определяет порядок аминокислот в полипептидной цепи и соответственно вид белка, который продуцируется. В результате этих исследований ген был определен как участок ДНК, содержащий основную последовательность для кодирования белка плюс коды начала и конца. Как мы уже упоминали раньше, такие простые взаимоотношения между последовательностью кодонов и последовательностью аминокислот, которые наблюдаются у прокариот, в целом не применимы для эукариот (к ним относятся и все многоклеточные организмы), у которых в молекуле ДНК имеются кодирующие (экзоны) и некодирующие (интроны) последовательности. Перед тем кaк кодирующая последовательность будет использована для синтеза белка, интроны должны быть удалены. Открытие кодирующих и некодирующих последовательностей у эукариот привело к тому, что предложенное ранее определение гена пришлось усложнять с учетом новых данных.
9.3.2. Транскрипция, трансляция и факторы, влияющие на эти процессы
Мы знаем, что ДНК находится в ядре клетки, а синтез белка происходит на рибосомах, находящихся в цитоплазме. Какие же молекулярные процессы имеют место при синтезе белка? Один из таких процессов называется транскрипцией и осуществляется с помощью другой нуклеиновой кислоты, которая носит название рибонуклеиновой (РНК). РНК представляет собой, так же как и ДНК, цепочку нуклеотидов, однако углеводным компонентом является рибоза, а одно из азотистых оснований ДНК - тимин - заменено на другое - урацил. В клетках эукариот транскрипцию осуществляет специальный фермент РНК-полимераза II. Молекула полимеразы прикрепляется к стартовой последовательности ДНК (промотору) и, двигаясь вдоль нити ДНК, начинает строить молекулу РНК по тому же принципу шаблона, который используется для дупликации ДНК. По достижении другого специфического участка ДНК - сигнала терминации - полимераза отделяется и высвобождает новосинтезированную молекулу РНК, которая носит название информационной, или матричной (мРНК). Эта одноцепочечная мРНК является копией ДНК и содержит код для последовательности аминокислот в белке, кодируемом данным участком ДНК. Затем мРНК перемещается из ядра в цитоплазму клетки к рибосомам, где на ее основе происходит построение полипептидных цепей, или трансляция. На ход транскрипции и трансляции могут влиять другие белки, относящиеся к регуляторным. Белки, влияющие на транскрипцию (факторы транскрипции), являются гетерогенной группой белков, которые либо путем активации, либо путем ингибирования регулируют экспрессию гена, то есть ту степень активности, с которой ген участвует в синтезе специфического для него белка. Факторы транскрипции определяют, будет ли, и в какой степени, ген экспрессирован. Для транскрипции у эукариот, в связи с существованием экзонов и нитронов, характерно включение в процесс дополнительного этапа редакции мРНК. Первоначальная транскрипция РНК включает считывание как кодирующих, так и некодирующих участков (формируется незрелая мРНК). Затем она должна быть трансформирована в зрелую мРНК, которая не имеет некодирующих фрагментов. Такая редакция осуществляется с помощью специального клеточного механизма, который носит название сплайсинга. Если что-то мешает сплайсингу, то незрелая мРНК становится нестабильной и исчезает из клетки. По количеству и разнообразию стабильной мРНК можно судить о разнообразии и степени активности генов, синтезирующих белки. Интересно, что сплайсинг может осуществляться различными путями, что позволяет на основе одной незрелой мРНК создавать различные формы зрелой. Это так называемый альтернативный сплайсинг. Альтернативный сплайсинг регулируется факторами клеточной среды. Примером альтернативно сплайсируемого гена является ген, который кодирует либо кальцитонин, пептид, включенный в регуляцию кальция, либо один из нейрогормонов. Если процессинг незрелой мРНК осуществляется в нейронах, то первичный транскрипт преобразуется в мРНК, кодирующую нейрогормон. А в клетках щитовидной железы альтернативный сплайсинг того же предшественника приводит к образованию молекулы мРНК, кодирующей пептид кальцитонин. Таким образом, специфика ткани регулирует тот конечный продукт, который синтезируется на основе одного и того же гена. Следовательно, возникающий в результате клеточный фенотип не определяется однозначно кодом ДНК, а слагается под влиянием многих частей клеточного механизма. Так что формула "один ген - один белок" для многоклеточных организмов оказалась неверной. Альтернативный сплайсинг приводит к увеличению разнообразия продуктов, которые могут быть закодированы в одном гене (см. рис. 9.3, а также тему 3).

Процесс трансляции
осуществляется на рибосоме.
В этом процессе участвует еще один вид
РНК - транспортная РНК (тРНК). Транспортная
РНК необходима, поскольку не существует
прямой комплементарности между мРНК и
аминокислотами. Транспортная РНК
является посредником между мРНК и
аминокислотами. В процессе сборки белка
различают три стадии - начало, продолжение
и окончание. Каждая из этих стадий также
может регулироваться.
После
трансляции полипептидные цепи складываются
в двух- и трехмерные структуры, образуя
белки различной конфигурации. Форма
зрелого белка определяет его функциональный
особенности. На форму и, следовательно,
на функции белка могут также влиять
различные процессы, происходящие в
клетке. Когда процесс синтеза белка
закончен, конечный продукт должен быть
транспортирован в определенное место
внутри или вне клетки, где он начнет
выполнять свою функцию. Этот процесс
также подвержен регуляции (рис.
9.3).
Среди
всех органов тела мозг занимает первое
место по числу активных генов. По
некоторым оценкам, число различных
молекул стабильной мРНК, продуцируемых
в мозге и специфичных для этого органа,
может быть около 150 000 (Changeux
J.P., 1997). Однако, несмотря
на такое многообразие экспрессирующихся
в мозге генов, количество синтезирующейся
в клетке мРНК
многих мозгоспецифичных генов относительно
невелико, транскрипция этих генов
осуществляется на низком уровне. Для
сравнения можно сказать, что в хрусталике
глаза продуцируется всего около 3000
различных мРНК, но количество каждой
из них достаточно велико, что говорит
о высокой интенсивности
транскрипции.
Интересно
оценить использует ли мозг, при таком
высоком количестве активных генов всю
ДНК, имеющуюся в хромосомах. Известно,
что в оплодотворенной яйцеклетке мыши
содержится 6x10-6 микрограмм ДНК. Если
произвольно порезать эту цепь на сегменты
(в соответствии с кодированием белка
молекулярным весом 40 000), то получится
примерно два миллиона таких сегментов.
Максимальное число генов не может быть
больше этого числа. Если учесть наличие
некодирующих последовательностей и
большого количества повторяющихся
последовательностей, то структурные
гены окажутся представлены лишь небольшой
частью всей ДНК. На сегодняшний момент
оценки максимального числа структурных
генов очень приблизительны и для мыши
составляют от 20 000 до 150 000. Последнее
число сопоставимо с разнообразием мРНК,
обнаруженным в мозге.
Оценить
количество генов, которые экспрессируются
в мозге и неактивны в других тканях
организма, можно экспериментально,
создавая тканеспецифичные кДНКовые
библиотеки. Первые работы по оценке
мозгоспецифичных генов были сделаны
на крысах. Было показано, что из
приблизительно 100 000 генов, которые
составляют весь геном крысы, около 60000
экспрессируются в клетках мозга, причем
около половины этих генов транскрибируются
только в мозге, а в других тканях эти
гены "молчат". Н. Жао с сотрудниками
провел анализ кДНК-библиотеки мозга
человека. Оказалось, что из 2505 случайно
выбранных рекомбинантных клонов
библиотеки около половины имеют
мозгоспецифическую экспрессию. Если
учесть, что в результате альтернативного
сплайсинга
количество мозгоспецифических
транскриптов еще более увеличивается,
то получается, что почти каждый второй
ген в геноме человека связан с обеспечением
функций нервной системы.
Таким
образом, процессы, происходящие при
считывании генетической информации с
ДНК и при последующем синтезе белка,
являются многоступенчатыми, и на каждой
ступени возможна регуляция. Это означает,
что между генотипом и фенотипом клетки
не существует однозначного соответствия,
а конечный результат целой цепи
превращений зависит от динамических
событий внутри и вне клетки. Регуляция
определяет, будет ли ген экспрессирован,
когда, где и в какой степени, какой белок
будет синтезирован на основе данного
гена, как он будет модифицирован и где
будет локализован. Регуляция может
происходить на любой ступени экспрессии
гена, но в основном она происходит при
процессах транскрипции. Здесь определяется,
будет ли вообще осуществляться
транскрипция, и если да, то с какой
скоростью она будет происходить и какова
будет ее продолжительность. Таким
образом, большинство регуляторов - это
факторы транскрипции.

Из всех видов тканей наибольшее разнообразие клеток и клеточных продуктов представлено в нервной системе. Это может обеспечиваться двумя путями: либо каждая клетка экспрессирует лишь небольшую часть своего генома, либо она использует один и тот же набор генов различным образом (вспомним альтернативный сплайсинг). Возможно, в нервной системе используются оба этих механизма, каждый из которых требует высоких уровней регуляции . Некоторая часть процессов регуляции связана с формой самой хромосомы. Например, для млекопитающих характерно явление, которое называется инактивацией одной из двух X-хромосом у представительниц генетически женского пола. Оказывается, что одна из Х-хромосом в клетках тела женской особи плотно скручивается и превращается в так называемое тельце Барра. В таком видоизмененном виде очень мало генов X-хромосомы оказываются доступными транскрипции, и она функционально напоминает более мелкую Y-хромосому. Такая инактивация осуществляется случайным образом и, если аллели обеих Х-хромосом различаются, может возникнуть явление мозаичности, известное, например, для кошек, имеющих черепаховую расцветку. Регуляция может быть также связана с изменениями в хроматине. Дело в том, что молекула ДНК не существует сама по себе, а находится в структуре, именуемой хроматином. Эта структура включает также белки - гистонные комплексы, и свыше 90% ДНК обычно обернуто вокруг гистонных комплексов, образуя четкоподобные структуры, называемые нуклеосомами (примерно по 150-200 пар оснований в каждой нуклеосоме) (рис. 9.4). Местоположение нуклеосом также может регулировать транскрипцию.
Большинство факторов транскрипции также являются белками, то есть продуктами трансляции. Это означает, что продукт, закодированный одним геном, может влиять на транскрипцию других генов, а иногда и того же самого гена. Следовательно, активность одной части генома может влиять на активность других его частей. Некоторые факторы транскрипции, экспрессированные генами, могут иметь широкое влияние на активность генома. Наибольшее внимание в этом отношении привлекают так называемые ранние гены (в английском варианте immediate early genes) и экспрессируемые ими продукты.
9.3.3. Ранние гены и их роль в развитии
Впервые ранние гены были идентифицированы в культуре ткани при исследовании механизмов активации генома в ответ на добавление в среду факторов роста, приводящих к клеточной пролиферации (увеличению массы клеток путем митоза) и дифференцировке. Эти исследования привели к открытию класса генов, которые активируются в течение нескольких минут после воздействия на клетки факторов роста, причем эта транскрипционная индукция происходит даже в том случае, если в среду добавлены ингибиторы синтеза белка. Оказалось, что эти гены кодируют ядерные белки, которые являются транскрипционными факторами и регулируют экспрессию других генов. Одними из первых в семействе ранних генов были идентифицированы гены c-fos и с-тус. Оказалось, что ранние гены широко распространены в живых организмах, причем один и тот же ген встречается у далеких в эволюционном отношении организмов. Это указывает на их важную биологическую функцию в нормальных клетках. Было показано, что ранние гены в условиях нормального функционирования характеризуются быстрой, но преходящей экспрессией в ответ на различные сигналы из внеклеточного пространства. Они способны реагировать на нейротрансмиттеры (передатчики нервных импульсов), на различные белковые факторы роста (например, фактор роста нервов - см. далее). Таким образом, действие разнообразных экстраклеточных сигналов приводит к запуску в клетках систем вторичных мессенджеров (посредников) и активации различных транскрипционных факторов, способных взаимодействовать друг с другом и активировать синтез мРНК генов-мишеней, обеспечивая тем самым специфичность и разнообразие восприятия клеткой различных сигналов. Считается, что ранние гены и их продукты могут быть одним из механизмов, посредством которых осуществляется координация между онтогенетическими событиями в нервной системе и во внешней среде. Посредством участия ранних генов некоторая генетическая активность может концентрироваться в определенных клетках и координироваться с другими процессами, происходящими в нервной системе или во внешней среде. Исследования показывают, что ранний опыт может значительно влиять на экспрессию ранних генов в клетках коры головного мозга. Так, К.М. Розен (1992) измерял уровень содержания мРНК ранних генов в зрительной и фронтальной коре котят, воспитывавшихся в темноте. Оказалось, что даже короткая световая экспозиция (всего один час) драматически увеличивает активность ранних генов в зрительной, но не во фронтальной коре. Примечательно, что эта активность была выше, если световая экспозиция приходилась на более ранний период развития (5-недельные котята в сравнении с 20-недельными). Таким образом, опыт может регулировать молекулярную генетическую активность. Накопленные данные о механизмах экспрессии ранних генов позволили предположить, что они могут не только играть важную роль в развитии нервной системы, но и быть связанными с такими важными процессами, как обучение и консолидация памяти (то есть процесс перехода информации из кратковременной памяти в долговременную). К настоящему моменту накоплено очень много данных об экспрессии ранних генов в мозге животных при обучении (см. например: Анохин К.В., Судаков К.В., 1993). Обобщая полученные результаты, можно выделить несколько основных свойств экспрессии ранних генов в мозге при обучении. В обычном состоянии, при отсутствии во внешней среде значимых раздражителей, экспрессия ранних генов в мозге животных осуществляется на базальном уровне с очень слабой интенсивностью. При помещении животных в новую среду или обучении их новым навыкам происходит очень быстрое и значительное усиление экспрессии ранних генов, причем наблюдаемая индукция транскрипции достигает своего максимума через 15-30 минут после начала воздействия, а уже через 60-90 минут уровень мРНК ранних генов возвращается к первоначальному базовому значению. Соответственно максимальный синтез белков, кодируемых этими генами, детектируется через 1-2 часа после воздействия, и через 4-5 часов индукция завершается (Там же). Примечателен тот факт, что по мере выработки и автоматизации нового навыка уровень экспрессии генов c-fos и c-jun постепенно снижается, а выполнение уже приобретенного навыка не вызывает индукции ранних генов. Привыкание к процедуре, вызывающей у животного стресс, также приводит к значительному снижению экспрессии ранних генов. При замене некоторых объектов во внешней среде или частичном изменении некоторых параметров условного сигнала затухавшая по мере обучения экспрессия ранних генов вновь достигает почти первоначального уровня (Там же). Эти данные послужили основой для гипотезы о том, что активация в мозге ранних генов происходит в том случае, когда животное сталкивается с новыми для него ситуациями и возникающая при этом экспрессия ранних генов отражает субъективную оценку животным новизны (Там же). В последние годы были получены прямые доказательства участия ранних генов в механизмах обучения и формирования памяти. Современные методы молекулярной биологии позволяют специфично и обратимо блокировать экспрессию отдельных генов. Этого можно добиться, вводя в клетки короткие последовательности олигонуклеотидов, комплементарных участку мРНК интересующего гена. Связавшись с мРНК, такие антисмысловые (антисенс-) последовательности блокируют трансляцию, и в клетке не синтезируется белок данного гена. В экспериментах на крысах и цыплятах было показано, что введение в критические для запоминания области мозга антисенс-олиго-нуклеотидов, временно "выключающих" экспрессию гена c-fos, вызывает у обученных животных амнезию. Эти данные свидетельствуют о том, что ранние гены очень чувствительно реагируют на изменения во внешней среде и являют собой критическое звено в механизме воздействия среды и приобретаемого в результате этого индивидуального опыта над геном нервных клеток. Таким образом, благодаря ранним генам даже краткие средовые сигналы могут быть преобразованы в каскад продолжительных событий, приводящих к изменениям внутри клеток и к межклеточным взаимодействиям. Тем самым ранние гены могут участвовать как в закладке нервной системы в эмбриогенезе, так и в ее пластических изменениях после рождения. Помимо ранних генов, в регуляцию генной активности и межклеточного взаимодействия включены и другие продукты. Одними из них являются гормоны.
9.3.4. Гормоны и их роль в генетической регуляции
Гормоны являются одним из биологически активных соединений, которые также способны принимать участие в регуляции дифференциации и активности нервных клеток на генетическом уровне. Стероидные гормоны (эстроген, тестостерон, прогестерон и некоторые другие) являются жирорастворимыми молекулами, благодаря чему они могут свободно проникать через клеточные мембраны в цитоплазму и ядро клетки. Для регуляции клеточной активности гормон должен не просто проникнуть внутрь клетки, но еще связаться со специфическим внутриклеточным рецептором. Рецептор представляет собой молекулярный комплекс, имеющий область связывания с ДНК, область связывания с гормоном и область, отвечающую за регуляцию транскрипции. После связывания гормона со своим рецептором результирующий комплекс способен прикрепиться к акцепторным участкам на хроматине и регулировать транскрипцию. Регуляция может выражаться в том, будет ли происходить транскрипция, какова будет ее скорость и с какого участка, или сайта (сайту соответствует участок ДНК, представляющий одну пару нуклеотидов), она начнется. Хотя стероидные гормоны и могут проникать во все клетки, но не все клетки, а только клетки-мишени имеют для них соответствующие рецепторы. Гормоны могут регулировать и свою собственную активность посредством процессов с обратной связью, которые регулируют число рецепторов в клетках-мишенях в сторону понижения или повышения. И у мыши, и у человека известны мутации, которые затрагивают структурные гены молекулярных рецепторов гормонов. Если такие мутации имеют место, то все ткани, включая мозг, становятся невосприимчивыми к действию соответствующего гормона. На продукцию и секрецию гормонов значительно влияет средовая стимуляция. Поэтому на генетическую активность в клетках-мишенях может влиять средовой опыт, который приводит к изменению содержания гормонов в организме. Это еще один путь влияния внешней среды на активность генов в развитии. Особенно интенсивно изучается влияние тестостерона на развитие нервной системы и поведения. Помимо специфических гормональных рецепторов, нейроны имеют еще один вид рецепторов, которые находятся на поверхности клеток и могут присоединять к себе гормоны, нейротрансмиттеры и нейромодуляторы - это так называемые G-белки.
9.3.5. Регуляторная роль g-белков
Многие гормоны, нейротрансмиттеры и нейромодуляторы влияют на активность нейронов с помощью специальных белковых рецепторных комплексов, имеющихся на поверхности клетки. Эти рецепторы были названы G-белками, поскольку они соединяются с внутриклеточным гуанозин-три-фосфатом (ГТФ). G-белки состоят из трех связанных с мембраной белков, которые называются альфа-, бета- и гамма-субкомпонентами. Когда рецептор активируется, альфа-субкомпонент связывается с ГТФ и отделяется от остальных субкомпонентов. Как альфа, так и бета-, гамма-субкомпоненты могут служить триггерами (то есть пусковыми механизмами) целых каскадов клеточных событий. Активированные G-белки способны влиять на проницаемость мембранных каналов, стимулировать высвобождение Са2+ из клеточных резервов, который, в свою очередь, может активировать Са2+- зависимые ферменты и запускать целый ряд других клеточных событий, которые могут непосредственно влиять на транскрипцию, трансляцию и посттрансляционные клеточные события в нейроне. Активированные G-белки имеют продолжительность жизни от секунд до минут. Затем происходит самоинактивация G-альфа, после чего он реаггрегирует с бета- и гамма-субкомпонентами и возвращается в исходное состояние. Таким образом, G-белки - это еще один путь влияния средовых событий на транскрипцию, трансляцию и посттрансляционные изменения в нервной клетке, который открывает возможности для влияния раннего опыта на структуру и функции нервной ткани. Мы специально довольно подробно остановились на некоторых аспектах современной молекулярной генетики, чтобы показать, насколько велики возможности регуляции генетической активности нервных клеток, причем не только со стороны внутриклеточных агентов, но и таких событий в межклеточных взаимодействиях, которые непосредственно связаны со средой вне организма. Внешние средовые факторы могут влиять на активность ДНК и клетки в целом только через внутреннюю химическую среду организма. Это происходит благодаря существованию специальных, созданных эволюцией приспособлений, которые способны трансформировать факторы внешней среды в биохимические субстанции. Это специальные биологические преобразователи, которые превращают энергию внешней среды в химические компоненты. В нервной системе такие преобразователи представлены наиболее широко. Вспомним рецепторы и те события, которые происходят при возникновении и передаче нервных импульсов. Отсюда понятно, что говорить о какой-либо жесткой генетической запрограммированности развития нервной системы, а следовательно и поведения, не приходится. В различные периоды развития события во внешней среде могут вмешиваться, изменяя работу генетического аппарата, включая одни гены и выключая другие, или изменяя интенсивность их экспрессии (см. также Хрестомат. 9.1).
9.4. Морфогенез нервной системы и факторы, влияющие на этот процесс
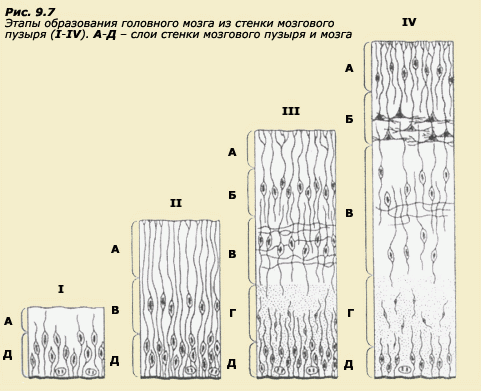
Не нужно доказывать, что особенности поведения теснейшим образом связаны со структурной и функциональной организацией нервной системы. Поэтому раннее развитие, в том числе и эмбриональное, когда происходит дифференциация нервных клеток и образование связей между ними и прочими системами организма, является одним из важнейших этапов, на котором формируются многие индивидуальные особенности поведения человека. Что же известно об этом современной науке? Центральная нервная система состоит из огромного числа клеток, отличающихся высочайшей степенью дифференциации по сравнению со всеми другими тканями организма. Эти клетки образуют сложные функционально значимые связи между собой и с остальными органами и тканями. Клетки образуют скопления, популяции, обладающие специфической морфологией и функциями. То есть по мере развития складывается сложнейшая функциональная система организма. И вместе с тем все клетки, слагающие эту систему, обладают одним и тем же набором генов. Мы уже упоминали, что количество ДНК в клеточном ядре мыши составляет 6х10-6 микрограмм. Это число для дрозофилы в 25 раз меньше. Однако если мы захотим сравнить в этом отношении мышь и человека, то получим поразительный результат. Оказывается, количество ДНК приблизительно одинаково у мыши, шимпанзе и человека. Это кажется парадоксом, если учесть, насколько возрастает сложность организации мозга при переходе от мыши к человеку. Число мозговых клеток увеличивается с 5-6 миллионов до нескольких десятков миллиардов. Вместе с тем количество ДНК в ядре оплодотворенной клетки существенно не меняется. Еще больше поражает то, что максимальное количество генов на такое количество ДНК (пусть это будет даже 200 000 или миллион) абсолютно несопоставимо с разнообразием нейронов, не говоря уже о синапсах. Таким образом, эволюционное усложнение мозга не сопровождается, по всей видимости, параллельным усложнением организации генома. Какой же механизм позволяет сформировать сложнейший орган человека - его мозг - на основе столь малого числа генетических детерминант, как происходит дифференциация нейронов, как складываются их популяции, как они образуют функционально значимые связи между собой и с иннервируемыми органами? На все эти вопросы пытается ответить интенсивно развивающаяся область науки - нейробиология развития, однако она еще очень далека от того, чтобы дать полное объяснение процессам морфогенеза нервной системы. Анатомические изменения в нервной системе координированы с подобными изменениями в остальном теле и с появлением функции. Эта координация осуществляется путем реципрокного взаимодействия между растущей нервной системой и остальными органами и тканями. В результате создается соответствие между структурой и функцией. В основе развития нервной системы лежат динамические процессы, идущие при участии внутренних и внешних по отношению к нейрону или нейронной популяции событий. События, связанные с развитием отдельной клетки, включают в себя рождение клетки, миграцию, дифференциацию, образование связей и возможную гибель на ранних этапах онтогенеза. Рождение. Увеличение числа нейронов происходит в результате митоза, который приводит к появлению новых клеток из уже существующих. У позвоночных большинство нейронов рождается на ранних стадиях развития в результате деления клеток неврального эпителия, который тянется вдоль нервной трубки. На 25-й день эмбриона человека нервная трубка еще открыта с обоих концов, но позже она замыкается, а в передней ее части образуются три пузырька, которые будут формировать головной мозг. Между 4 и 6 неделями жизни нервная система эмбриона человека напоминает нервную систему рыбы. Митотическое деление нервных клеток в этот период происходит настолько интенсивно, что через несколько месяцев их количество уже составляет несколько десятков миллиардов. Временами скорость деления достигает 250 000 клеток в минуту. Через 16-20 недель после оплодотворения деление нервных клеток, по-видимому, прекращается. Таким образом, количество нервных клеток достигает своего максимума задолго до рождения. Образование новых нейронов возможно и в постэмбриональный период, но это не является правилом. Конечное же число нейронов определяется не только процессами их рождения, но и процессами естественной гибели, которые являются важным элементом развития нервной ткани. Естественная гибель нейронов играет особенно существенную роль в морфогенезе нервной ткани у птиц и млекопитающих. По мере роста, дифференцировки и установления функциональных связей сверхпроизведенные нейроны, которые как бы не находят себе места в складывающейся функциональной системе, погибают. Гибель нейронов регулируется локальными условиями и является относительно поздним эмбриональным событием, обычно происходящим после формирования синапсов. В промежутке между своим рождением и решающим моментом, "жить ему или погибнуть", нейрон мигрирует к месту своей окончательной локализации, проходит дифференциацию и устанавливает свои первичные афферентные и эфферентные связи (рис. 9.5 а, б). Миграция. Большинство нейронов имеют место своей локализации на некотором расстоянии от места своего рождения. Так что миграция нейронов является обязательным и важным этапом нейронального развития. До сих пор является загадкой, как нейроны находят место своего окончательного назначения. Очевидно, главным источником информации по поводу миграции являются поверхности, окружающие клетку. Эти поверхности отличаются по своей адгезивности (то есть вязкости, способности склеивать клетки между собой), и именно это свойство, по-видимому, является наиболее важным для установления путей миграции. Градиент адгезивности заставляет нейрон передвигаться. Большая часть адгезивного субстрата для нейронов связана с другим важным типом клеток, представленных в нервной ткани, а именно с клетками глии. Например, при формировании коры большую роль играет радиальная глия, которая простирает отростки сквозь все развивающиеся слои коры вплоть до ее поверхности. По этим "дорогам" нейроны, образующиеся в глубинных слоях вблизи мозгового желудочка, скользят в более поверхностные слои коры (рис. 9.6 а, б). Однако существуют и другие "маршруты" для передвижения нейронов. Известно, что клетки коры мозга образуют шесть слоев. V взрослого слой I является самым поверхностным, а слой VI - самым глубоким. Зона деления нервных клеток находится еще глубже. Было бы естественно предположить, что по мере деления слои клеток накапливаются подобно осадочным породам, и более молодые клетки оказываются ближе к зоне деления. На самом деле случается обратное. Слой VI - самый глубокий - соответствует наиболее рано сформировавшимся клеткам, тогда как все остальные слои занимают свои места, проходя через все уже установившиеся слои, так что слой I соответствует самым молодым клеткам (рис. 9.7). Принципы умножения, миграции и размещения клеток в слоях, по-видимому, являются общими для всей коры, что позволяет предположить участие в этом процессе небольшого числа генов.
Дифференциация. Другой загадкой нейроэмбриогенеза являются процессы дифференциации нейронов. Как клетки с идентичным генным составом преобразуются в столь разнообразные формы, которые характерны для нейронов зрелого мозга? Во-первых, клетки нервной ткани дифференцируются на два крупных класса. Клетка становится либо нейрональной, либо глиальной. Внутри каждого из этих классов также идет дифференцировка, особенно характерная для нейронов. Нейроны отличаются по форме, по размерам, по характеру своих отростков и по нейротрансмиттерам, которые они используют (адренэргические, холинэргические). Причины возникновения этих различий до конца не поняты. Ясно только, что они включают как процессы взаимодействия клетки с ее окружением, так и дифференциальную активацию и подавление генов на определенных стадиях развития. Проходя последовательные стадии развития, нейрон приобретает свойства, которые могут влиять на прохождение последующих стадий. Скорее всего, решающие воздействия на окончательное формирование нейрона оказывают процессы взаимодействия с другими клетками. Например, нейротрансмиттерная специфичность нейронов в автономной нервной системе зависит от клеточного окружения. Вещества, продуцируемые теми тканями, которые иннервирует данный нейрон, способны изменять его специфичность. Например, вещества из клеток сердца побуждают нейрон становиться холинэргическим. Выживание и гибель. Для выживания нейронов, по-видимому, решающим является воздействие трофических факторов. К трофическим факторам относят вещества, продуцируемые другими клетками, которые способны поддерживать рост, функционирование и выживание данной клетки. Число нейронов, образующихся в результате митоза на ранних эмбриональных стадиях, примерно в два или три раза превышает то количество нейронов, которое выживает на более поздних стадиях. Естественная нейрональная гибель происходит вскоре за установлением афферентных и эфферентных связей с другими клетками. Предполагается, что естественная гибель нейронов происходит в процессе конкуренции за ограниченный ресурс трофических факторов, поставляемых теми тканями, с которыми нейроны устанавливают функциональные связи. Однако точная природа этой конкуренции пока неясна. Нейроны, которые успешно взаимодействуют с клетками-мишенями, получают необходимые трофические факторы, а остальные, которые не могут успешно взаимодействовать, отмирают. Идет своего рода естественный отбор. Наиболее известным из трофических факторов является фактор роста нервов (ФРН), который был описан Р. Леви-Монтальчини (Levi-Montalcini R.) в 1975 г. Оказалось, что слюнная железа мыши является превосходным источником ФРН, что позволило вести интенсивные исследования с его применением. ФРН продуцируется естественным путем внутри нервной системы и ее периферических мишеней. Некоторым популяциям нейронов ФРН требуется для выживания и роста. Инъекция ФРН в развивающийся эмбрион увеличивает выживаемость нейронов в период естественной гибели клеток. Образование связей. Аксоны и дендриты корковых нейронов начинают появляться намного раньше, чем устанавливаются все шесть слоев коры. В это же время формируются и первые синапсы. Так, предшественники пирамидных клеток посылают свои аксоны к соответствующим ядрам таламуса, а аксоны таламических нейронов растут по направлению к коре. Связь между корой и таламусом начинает формироваться реципрокно и синхронно еще до того, как образуются II и III слои. Главные пути связей в мозговой коре человека закладываются еще до рождения. Многие процессы, лежащие в основе миграции нейронов, имеют место и при образовании отростков нервных клеток - аксонов и дендритов. Аксоны и дендриты движутся по направлению к клеткам-мишеням, постепенно отрастая из тела клетки. Скорость роста может составлять 15-20 микрон в час. На ведущем конце нейрита (аксона или дендрита) имеется специальная структура, называемая конусом роста (рис. 9.8).
Впервые эта
структура была описана С.
Рамон-и-Кохалем в 1890 г. Конус
роста реагирует на сигналы из
экстраклеточной среды, в том числе на
ФРН, который может привлекать конус
роста, и некоторые адгезивные
белки. Трофические
факторы, получаемые из локальной
клеточной среды, являются важными
регуляторами процесса образования,
роста и ветвления аксонов и дендритов.
Обычно начальная стадия формирования
нейрита отличается обильным ростом
ответвлений, часть которых будет в
дальнейшем элиминирована. Например,
мозолистое тело - проводящий путь из
одного полушария в другое - в неонатальный
период испытывает значительную потерю
аксонов, что связывается с реорганизацией
паттерна проекций корковых нейронов в
противоположное полушарие. Предполагается,
что подобные реконструкции являются
окончательным результатом клеточных
взаимодействий. Может быть, связи,
которые являются функциональными,
остаются, а остальные исчезают. Однако
имеется возможность для изменения
паттерна связи в течение жизни.
Как
только конус роста достигает конечной
цели, находит соответствующую
клетку-мишень, например, мышечную,
процессы роста тут же прекращаются, и
конус роста превращается в нервное
окончание, на котором постепенно
формируется зрелый синапс (рис. 9.8).
Каждая категория нервных клеток имеет
свою категорию мишеней, которую распознают
растущие нейриты, устанавливая связи
только с клетками соответствующего
типа. Таким образом, поведение конуса
роста детерминировано, по-видимому,
лишь немногими жестко запрограммированными
правилами, включая амебоидные движения
и узнавание своей мишени. Остальное
определяется случайностями развития,
что приводит к высокой вариабельности
морфологии нервных отростков.
 Интересно,
что этап онтогенеза,
на который приходится воздействие
трофического
фактора, накладывает
отпечаток на характер роста нервных
окончаний. Например, число первичных
дендритов, растущих прямо из тела
нейрона, может быть увеличено подачей
ФРН только в эмбриональный период, тогда
как применение ФРН на более поздних
стадиях ведет к росту ответвлений более
высокого порядка и не меняет число
первичных отростков. Таким образом,
влияние трофических веществ на клетку
зависит от степени дифференцированности
клетки. Наиболее подвержены изменениям
ветвления более высокого порядка, их
длина, а также такие морфологические
свойства, как число шипиков.
Гормоны
также могут оказывать трофические
влияния на развивающиеся нейроны,
которые имеют соответствующие рецепторы
для этих гормонов. И наконец, что для
нас особенно важно, стимуляция из внешней
среды и обратная связь от поведения
индивида могут иметь существенное
влияние на формирование связей в нервной
системе. Это означает, что изменения в
протяженности и ветвлении дендритов,
образовании синапсов
и ветвлении аксонов испытывают влияния
раннего опыта, то есть средовые влияния.
Многие из этих влияний действенны во
время позднего эмбрионального или
раннего неонатального развития, но
имеют место и в более поздние периоды
онтогенеза.
Таким
образом, нервная система создается
преимущественно в период эмбриональной
жизни из клеток с одинаковым генетическим
потенциалом, которые делятся, мигрируют,
дифференцируются, образуют связи и
выживают или погибают в период естественной
гибели клеток. Нервная система способна
перестраивать связи в течение жизни
под влиянием опыта. Синапсы могут
исчезать и появляться, а аксоны и дендриты
способны образовывать и убирать свои
отростки в течение своей жизни. Это
является отражением пластичности
нервной системы. Непрерывно происходящие
с
Интересно,
что этап онтогенеза,
на который приходится воздействие
трофического
фактора, накладывает
отпечаток на характер роста нервных
окончаний. Например, число первичных
дендритов, растущих прямо из тела
нейрона, может быть увеличено подачей
ФРН только в эмбриональный период, тогда
как применение ФРН на более поздних
стадиях ведет к росту ответвлений более
высокого порядка и не меняет число
первичных отростков. Таким образом,
влияние трофических веществ на клетку
зависит от степени дифференцированности
клетки. Наиболее подвержены изменениям
ветвления более высокого порядка, их
длина, а также такие морфологические
свойства, как число шипиков.
Гормоны
также могут оказывать трофические
влияния на развивающиеся нейроны,
которые имеют соответствующие рецепторы
для этих гормонов. И наконец, что для
нас особенно важно, стимуляция из внешней
среды и обратная связь от поведения
индивида могут иметь существенное
влияние на формирование связей в нервной
системе. Это означает, что изменения в
протяженности и ветвлении дендритов,
образовании синапсов
и ветвлении аксонов испытывают влияния
раннего опыта, то есть средовые влияния.
Многие из этих влияний действенны во
время позднего эмбрионального или
раннего неонатального развития, но
имеют место и в более поздние периоды
онтогенеза.
Таким
образом, нервная система создается
преимущественно в период эмбриональной
жизни из клеток с одинаковым генетическим
потенциалом, которые делятся, мигрируют,
дифференцируются, образуют связи и
выживают или погибают в период естественной
гибели клеток. Нервная система способна
перестраивать связи в течение жизни
под влиянием опыта. Синапсы могут
исчезать и появляться, а аксоны и дендриты
способны образовывать и убирать свои
отростки в течение своей жизни. Это
является отражением пластичности
нервной системы. Непрерывно происходящие
с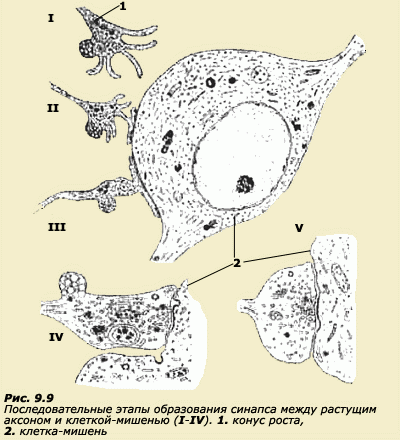 труктурные
и функциональные изменения в нервной
системе отражают события, происходящие
в остальных частях организма и во внешней
среде. В результате возникает адаптивное
соответствие между нервной системой и
другими частями организма, в особенности
сенсорной и двигательной системами,
через которые осуществляется взаимодействие
с внешней средой.
Поэтому
и повреждения мозга, возникающие в
ранние периоды онтогенеза, по всей
видимости, вызывают изменения,
принципиально отличные от тех, что имеют
место у взрослых. И это отличие обусловлено
не просто разным уровнем пластичности.
Вероятнее всего, ответом на раннее
повреждение может быть установление
необычных нервных связей путем
компенсаторного аксонального роста,
разрастания дендритов
и образования синапсов. В результате
происходят изменения в функциональной
и структурной организации оставшейся
ткани. Происходит как бы формирование
нового мозга, обладающего иной
функциональной организацией.
Небольшой
экскурс в нейроэмбриологию показывает,
что на каждом этапе развития и
морфологические, и функциональные
особенности организма являются
результатом взаимодействия между
множеством переменных. К ним относятся
все типы генов, в том числе и регуляторные,
различные физико-химические процессы,
происходящие внутри организма, и внешние
события, вызывающие целые каскады
онтогенетических превращений. Невозможно
понять отношения между организмом и
теми причинами, благодаря которым он
формируется, не привлекая теорию
динамических систем, ибо развивающийся
организм, безусловно, относится к
системам этой категории. Для познания
сложных и запутанных механизмов развития
наиболее перспективными являются
междисциплинарные исследования,
объединяющие подходы таких наук, как
генетика, нейрофизиология, эмбриология,
биология и психология развития Хрестомат.
9.2,
9.3).
труктурные
и функциональные изменения в нервной
системе отражают события, происходящие
в остальных частях организма и во внешней
среде. В результате возникает адаптивное
соответствие между нервной системой и
другими частями организма, в особенности
сенсорной и двигательной системами,
через которые осуществляется взаимодействие
с внешней средой.
Поэтому
и повреждения мозга, возникающие в
ранние периоды онтогенеза, по всей
видимости, вызывают изменения,
принципиально отличные от тех, что имеют
место у взрослых. И это отличие обусловлено
не просто разным уровнем пластичности.
Вероятнее всего, ответом на раннее
повреждение может быть установление
необычных нервных связей путем
компенсаторного аксонального роста,
разрастания дендритов
и образования синапсов. В результате
происходят изменения в функциональной
и структурной организации оставшейся
ткани. Происходит как бы формирование
нового мозга, обладающего иной
функциональной организацией.
Небольшой
экскурс в нейроэмбриологию показывает,
что на каждом этапе развития и
морфологические, и функциональные
особенности организма являются
результатом взаимодействия между
множеством переменных. К ним относятся
все типы генов, в том числе и регуляторные,
различные физико-химические процессы,
происходящие внутри организма, и внешние
события, вызывающие целые каскады
онтогенетических превращений. Невозможно
понять отношения между организмом и
теми причинами, благодаря которым он
формируется, не привлекая теорию
динамических систем, ибо развивающийся
организм, безусловно, относится к
системам этой категории. Для познания
сложных и запутанных механизмов развития
наиболее перспективными являются
междисциплинарные исследования,
объединяющие подходы таких наук, как
генетика, нейрофизиология, эмбриология,
биология и психология развития Хрестомат.
9.2,
9.3).
9.5. Роль эмбрионального и неонатального опыта в развитии поведения
Развитие является непрерывным процессом. В этом смысле рождение, то есть переход из внутриутробной среды во внеутробную, является лишь условной точкой отсчета, несмотря на всю радикальность такого события, как рождение. Непрерывность развития подтверждается тем, что неонатальное поведение младенцу поначалу во многом напоминает поведение плода внутри матки. Так, поза новорожденного представляет собой продолжение той позы, которая была приобретена в матке в последний триместр беременности. Для млекопитающих, эмбриональное развитие которых происходит внутри матки, наблюдение за эмбриональным периодом развития поведения, к сожалению, весьма ограничено из-за множества технических трудностей. Гораздо больше информации имеется на настоящий момент об эмбриональном поведении птиц. Однако использование приборов внешнего мониторинга и ультразвуковых устройств, а также разработка новых методов выхаживания очень незрелых недоношенных детей в последнее время расширили наши знания даже о самых ранних этапах развития поведения человека. Известно, что движения плода начинаются очень рано. Сердцебиения возникают примерно на 3-4 неделе после оплодотворения, первые спонтанные движения туловища и конечностей - на 10-й неделе, но мать начинает ощущать их примерно на семь недель позже. Какую же функциональную роль выполняют эти движения, полезны ли они для дальнейшей жизни или являются побочным продуктом развития двигательной системы? Контролируются ли они эндогенно или являются реакцией на стимуляцию? Результаты работ последних лет показывают, что некоторые особенности двигательной активности являются реакцией на специфику внутриутробной или ранней постнатальной среды. Оказалось, что плоды крыс способны отвечать на внешнюю стимуляцию, а нервная система плода способна перерабатывать тактильную и проприоцептивную сенсорную информацию. Следовательно, сенсорная обратная связь от движений, ограниченных маткой, может модифицировать нервную систему и управлять двигательной активностью. Учитывая чувствительность нервной системы к влиянию сенсорного опыта, можно предполагать, что ее модификация (через обратную связь) может вызывать изменения, сохраняющиеся и после рождения. Спонтанная активность плода человека является сложно организованной деятельностью, в которой представлены различные комплексы движений, однако окончательное значение ее не ясно. Например, у плодов часто наблюдается сопряженная активность головы и кисти и сосание большого пальца. Возможно, это облегчает постнатальную координацию этой деятельности, характерной для младенцев, находящихся в состоянии дискомфорта. Наблюдаются также эпизодические спонтанные дыхательные движения, которые могут быть важными для адекватной дифференциации ткани легких и правильной иннервации межреберных мышц. Известно, что у плодов крыс отклоняющиеся от нормы связи межреберных нервов с несоответствующими сегментами мышц элиминируются во внутриутробном периоде с началом дыхательных движений. Повторяющийся ток амниотической жидкости в легкие и обратно также важен для развития легких и респирации. Глотательные движения и заглатывание амниотической жидкости могут служить как для сиюминутных нужд плода (регулирование объема амниотической жидкости и ее вязкости, получение важных элементов питания, гормонов и иммунных факторов), а также быть подготовкой для перехода к заглатыванию молока и сосанию. Протекание жидкости через вкусовые и обонятельные рецепторы обеспечивает химические стимулы, которые могут играть важную роль в идентификации пищи. Плод млекопитающих, несомненно, отвечает на химические (вкус, обоняние), тактильные и слуховые стимулы и может запоминать пренатальный опыт. Например, у крыс в естественных условиях запах материнских сосков имеет общие элементы с запахом амниотической жидкости. Если в последние дни беременности в амниотическую жидкость впрыскивать пахучий цитраль, то новорожденные крысята предпочитают сосать из обработанных цитралью сосков матери. Детеныши, которые не подвергались такому внутриутробному опыту, избегают пахнущих цитралью сосков. Сенсорные системы развиваются в последовательности, которая является общей для всех позвоночных. Например, у крысы, которая рождается незрелой, еще до рождения начинают функционировать обоняние, вкус, температурная, тактильная и вестибулярная чувствительность. У человека все сенсорные системы обретают функциональный статус до рождения. Так, слуховая система плода примерно на 6-м месяце беременности уже имеет основные характерные черты взрослого организма. Тактильная рецепция складывается еще раньше. Спонтанная активность сенсорных систем возникает задолго до рождения. Единственная стимуляция, которая не достигает плода и не играет заметной роли в развитии - это зрительная. Интересны наблюдения за слуховым опытом плода. Слышание голоса матери является обычным ранним опытом плода. Голос матери легко проникает через ее ткани и амниотическую жидкость. К тому же он сопровождается кинестетической и тактильной стимуляцией плода в результате движений диафрагмы матери при разговоре. Таким образом, речь матери дает плоду слуховой опыт в сочетании с вестибулярной и тактильной стимуляцией. Плод особенно восприимчив к тоническим оттенкам речи матери, которые несут информацию и об эмоциональном состоянии. Именно благодаря этим характеристикам двухдневные младенцы способны узнавать голос матери. На слуховую систему плода воздействуют многие звуки, генерируемые матерью. Таким образом, мать обеспечивает плоду разнообразный слуховой опыт, который очень важен для появления многих когнитивных, эмоциональных и социальных способностей новорожденного. Было показано, что новорожденные способны дифференцировать взрывные согласные звуки речи, такие, как "д" и "т". Эта способность слишком сложна, чтобы происходить из пренатального или постнатального опыта, поэтому многие интерпретируют ее как свидетельство врожденных нервных механизмов, настроенных на акустические свойства речи. Однако недавно (Seeback B.S. et al., 1994) была создана компьютерная программа нейронной сети для демонстрации того, что такое категорийное различение может возникать из пренатального слухового опыта. Авторы подвергали модель воздействию естественных речевых звуков, измененных в соответствии со звуками, доступными плоду, и в соответствии с ограничениями в переработке информации слуховой системой плода. Программа приобрела способность к категорийному различению звуков речи английского языка и, более того, оказалась способна использовать свое категорийное восприятие для детекции и различения взрывных согласных языка хинди. Эти результаты показывают, что пренатальный опыт, вполне вероятно, является одним из источников начального развития речевых навыков. В целом исследования эмбриологии поведения демонстрируют, что многие поведенческие паттерны закладываются в эмбриональный период и имеют непосредственное отношение к событиям, связанным с внутриутробным опытом плода. Предполагается, что часть поведения плода является адаптацией к внутриутробным условиям, тогда как многие элементы поведения служат подготовительным этапом для дальнейшего развития (Michel G.F., Moore C.L., 1995). Например, регистрация ЭЭГ у плодов за несколько недель до рождения позволяет зафиксировать изменения, характерные для переходов от периодов бодрствования ко сну, причем в ЭЭГ сна уже отмечаются эпизоды интенсивной активности, характерные для парадоксального сна. Интересные наблюдения связаны с характерной для поврежденных асимметрией позы, которая включает предпочитаемое направление ориентации головы. В положении на спине новорожденный поворачивает голову в одну сторону (для большинства младенцев чаще вправо, чем влево). Рука и нога на той стороне, куда повернуто лицо, обычно разогнуты, тогда как другие рука и нога согнуты. Рука со стороны лица более активна - как по разнообразию, так и по частоте движений. Сдвиг вправо в предпочитаемом положении головы новорожденного создает латеральное смещение для визуализации руки и, следовательно, для действий этой рукой. Это латеральное смещение в сенсомоторном опыте рук предсказывает последующее предпочтение в использовании руки при контакте с предметами в период от 6 до 18 месяцев. Вообще вопрос о происхождении полушарной специализации функций и асимметрии рук постоянно дискутируется в нейропсихологии и генетике поведения. Имеются гипотезы о наследственной природе этих асимметрий. Например, в соответствии с гипотезой М. Аннет (1978) феномен предпочтения руки у человека объясняется действием как генов, так и случайных факторов в развитии, при этом отчетливый сдвиг в сторону правшей в популяции в целом объясняется действием одного гена, который М. Аннетт называет геном правого сдвига. Однако, как показывают последние наблюдения, события раннего опыта, по-видимому, могут играть немаловажную роль в возникновении полушарной специализации функций. Ф. Превик (1991, цит. по: Michel G.F., Moore C.L., 1995) предполагает, что пренатальный вестибулярный опыт плода может оказывать влияние на возникновение неонатальной асимметрии позы. Известно, что положение плода меняется в течение беременности, но асимметричный характер человеческой матки в комбинации со специфической гравитацией плода создают условия, результатом которых будет положение плода, которое является более вероятным, чем все остальные (инвертированный плод со спиной, повернутой к левой стороне матери) (рис. 9.10). При этом обычное движение матери вперед будет создавать асимметричный вестибулярный опыт для плода, поскольку это движение будет по-разному сказываться на левом и правом отолитических органах плода. Результатом асимметрии в стимуляции может быть большая чувствительность левого отолитического органа, чем правого. Поскольку левый отолитический орган имеет проекцию в правое полушарие, он обеспечивает правому полушарию активацию, отличную от левого. Активация левого органа может иметь результатом моторную активацию мышц, которые поворачивают голову вправо и изгибают туловище. Это вносит вклад в появление асимметрии неонатальной позы, что, в свою очередь, влияет на развитие асимметрии в использовании руки у младенца. Вестибулярная система - одна из наиболее рано развивающихся сенсорных систем, и она является активной во время развития плода. Возможно, асимметричная активация этой системы создает предпосылки для возникновения различных функциональных асимметрий у человека. Поскольку для животных не характерно прямохождение, этот феномен у них не наблюдается. Ф. Превик отмечает также, что полушария отличаются и по характеру преобладающих нейротрансмиттеров в некоторых структурах: если для правого полушария, по всей видимости, более характерна норадренергическая и серотонинергическая иннервация, то для левого - допаминергическая и холинергическая. Возможно, асимметрия вестибулярного опыта ответственна и за асимметрию в этих главных нейротрансмиттерах. Таким образом, еще очень мало известно о влиянии самого раннего опыта, связанного с эмбриональным и неонатальным периодами жизни, однако некоторые наблюдения свидетельствуют о том, что множество средовых факторов могут накладывать отпечаток на формирование нервной системы и развитие поведения.
9.6. Родительские эффекты в развитии
Образование гамет, оплодотворение и эмбриональное развитие у млекопитающих происходят внутри организма родителей, поэтому мы вправе задать вопрос, не влияет ли физиологическое состояние родителей на эти процессы. Особенно важно для развития организма состояние всех физиологических систем матери. В целом можно сказать, что генотип ребенка (то есть состав генов) не испытывает влияния таких факторов, как возраст, количество предыдущих родов или состояние здоровья родителей. Нормальные процессы мейоза, сопровождающиеся рекомбинацией хромосом, по-видимому, не зависят от состояния организма родителей. Наиболее чувствительными к физиологическому состоянию родителей являются два генетических процесса - это мутации и кроссинговер. Не будем подробно останавливаться на мутационном процессе, поскольку это особая область генетики. Отметим лишь, что существуют различные мутагенные факторы (радиация, химические агенты), и, если они имели место в период гаметогенеза, оплодотворения и эмбрионального развития, могут возникать различные аномалии, причиной которых являются произошедшие мутации. Что же влияет на кроссинговер? Эксперименты на животных показывают, что частота кроссинговера зависит от возраста и состояния процессов метаболизма родителей. Если будет доказано то же самое для человека, то это означает, что дети, рожденные в периоды низкой частоты кроссинговера, должны иметь большее количество сцепленных блоков аллелей, доставшихся им с хромосомой одного из прародителей (дедушки или бабушки), тогда как в периоды более высокой частоты кроссинговера они будут нести смешанные признаки обоих прародителей, поскольку их хромосомы будут содержать участки, происходящие и от дедушки, и от бабушки (см. схему кроссинговера в теме 3). Не так давно в генетике было открыто явление, получившее название генетического импринтинга. Термин, возможно, не совсем удачный, так как он никакого отношения к поведенческому явлению импринтинга не имеет. Генетический импринтинг проявляется в разной экспрессии одного и того же гена в зависимости от того, получен ли этот ген от отца (через спермин) или от матери (через яйцеклетку). Если получить мышиный эмбрион, все хромосомы которого происходят от родителя одного пола, то оказывается, что возникают два типа аномалий в развитии. Если все хромосомы получены от отца, то возникают аномалии самого эмбриона; если все хромосомы получены от матери, то возникают нарушения в развитии плаценты. Типичным примером генетического импринтинга является время начала заболевания при хорее Гентингтона: если патологический аллель передан отцом, симптомы болезни появляются в юности, а если матерью, то не ранее 40 лет. Таким образом, гены, находящиеся в хромосомах, как бы несут на себе печать своего происхождения (из яйцеклетки или из сперматозоида), и для нормального развития нужны не просто нормальные гены, а обязательно гены, полученные от каждого из родителей. Механизм генетического импринтинга неясен. По-видимому, в процессе образования гамет происходит модификация генов, которая идет по-разному при гаметогенезе яйцеклеток и спермы. Геном, идущий от самки, будет иметь гены, модифицированные по паттерну яйцеклетки, а геном, идущий от самца, соответственно, по паттерну сперматозоида. Интересно, что эти модификации не передаются через поколения. Это означает, что все гены в гаметах самки, например, будут модифицированы по типу яйцеклетки независимо от того, в какой хромосоме они находятся - той, что унаследована от дедушки или той, что - от бабушки. Материнский эффект. Особое место в родительских влияниях на развитие занимает так называемый материнский эффект. Влияние матери на фенотип потомства обнаруживается гораздо чаще, чем влияние отца. Эти влияния связаны с цитоплазматическими эффектами яйцеклетки, внутриутробными событиями и постнатальными влияниями, связанными со вскармливанием и уходом за потомством. Все три типа влияний могут изучаться в экспериментах на животных. Чтобы отделить влияния внутриутробной среды от влияния яйцеклетки, самкам одной инбредной линии мышей или крыс до спаривания делается пересадка яйцеклетки от самок другой инбредной линии. Чтобы изучать влияние постнатальной материнской среды, обычно потомство одной самки передается для вскармливания и воспитания другой. Таким образом, путем комбинации различных ситуаций можно выяснить эффект каждого из трех типов материнских влияний. В экспериментах на мышах, например, было показано, что особенности постнатальных материнских влияний способны снижать агрессивность самцов, происходящих из генетически более агрессивной линии. В этих же экспериментах было выявлено, что на агрессивность в той или иной мере влияют все материнские факторы. Таким образом, один и тот же ген имеет разное фенотипическое проявление в поведении в зависимости от условий развития, определяемых материнскими факторами. Еще одно интересное наблюдение касается так называемого хэндлинга. Когда изучается влияние раннего опыта на поведение животных, часто используют процедуру хэндлинга. Она заключается в том, что детенышей мышей и крыс в течение первых трех недель после рождения регулярно берут в руки. Оказалось, что такие животные, будучи взрослыми, проявляют большую любознательность и эмоциональность, чем интактные, не получавшие хэндлинга, даже если хэндлинг ограничивался трехминутными эпизодами. Первоначально считалось, что изменение поведения животных является прямым следствием хэндлинга, однако последующие наблюдения за взаимоотношениями животных, подвергавшихся хэндлингу, с их матерями показало, что после хэндлинга и у мышей, и у крыс матери гораздо больше "тормошили" своих детенышей по сравнению с теми, которые хэндлингу не подвергались, так что изменения в поведении могли возникнуть и за счет изменений, возникших в поведении матерей экспериментальной группы. Интересны также наблюдения за развитием недоношенных детей, которых после рождения помещали в инкубатор. Оказалось, что дети, которых время от времени встряхивали и тормошили, быстрее набирали вес и были выписаны из больницы раньше контрольных. У человека, как и у других млекопитающих, тесная физиологическая близость матери и ребенка возникает в момент зачатия и сохраняется вплоть до отнятия от груди. При этом не только ребенок испытывает влияния со стороны физиологического и психического состояния матери, но и в организме и поведении матери происходят ответные изменения, связанные с вынашиванием, рождением, вскармливанием и воспитанием ребенка. Говорят, что мать и дитя находятся в реципрокных отношениях. Через мать осуществляется регуляция среды плода и новорожденного, которая обеспечивает поддержание систем гомеостаза. Внутриутробная среда поддерживает уровни температуры, кислорода, воды, электролитов и питательных веществ. После рождения наиболее универсальным регулирующим веществом является молоко матери, которое поставляет организму ребенка питательные вещества, электролиты, воду, гормоны и иммунные факторы. Помимо вскармливания, с материнским уходом связаны и другие регуляторные процессы. В первые же дни после рождения в результате реципрокных отношений у матери и ребенка устанавливается специфический режим взаимодействия. Мать и дитя приспосабливаются друг к другу. Такое установление взаимовлияний происходит уже в первые две недели жизни ребенка. Те дети, которые после рождения попадают в больничные условия (особенно если они находятся в инкубаторе, и матери не позволяется ухаживать за ребенком), в этот период не имеют тесных контактов с матерью, что неблагоприятно сказывается на их психическом развитии. Этого не происходит в тех случаях, когда мать допускается к уходу за ребенком. Интересны данные, полученные недавно в нашей стране на младенцах со зрительной депривацией в результате врожденной катаракты. Оказалось, что первые месяцы жизни являются критическими, когда весь комплекс взаимодействия с матерью имеет принципиально важное значение для дальнейшего развития эмоционального и социального поведения ребенка. Исключение из этого комплекса зрительного входа (улыбка матери, выражение ее лица при общении) в первое полугодие жизни, несмотря на то, что сохраняются слуховые и тактильные контакты, приводит в дальнейшем к необратимым изменениям в эмоциональном и социальном поведении ребенка, даже если катаракта снимается на первом году жизни (в среднем, в шесть-восемь месяцев), и ребенок начинает видеть. По-видимому, в первые месяцы жизни, существует критический период для формирования каких-то базовых характеристик, обеспечивающих в дальнейшем эмоциональное и социальное благополучие ребенка. Зрительная депривация в этом периоде приводит к необратимым изменениям в психике ребенка (Сергиенко Е.А. и др., 1993; Строганова Т.А. и др., 1995; Строганова Т.А., Цетлин М.Н., 1997). Таким образом, ранние реципрокные взаимодействия матери и ребенка имеют принципиальное значение для адекватного психологического развития ребенка. Межпоколенные влияния. Вообще системы родительской заботы на ранних этапах развития имеют некоторые стабильные особенности, которые формируют часть негенетической наследственности потомства. Некоторые из них являются видоспецифическими, другие присущи отдельным семьям (традиции, передаваемые из поколения в поколение). Например, молодые самцы снегиря, находясь в гнезде, перенимают пение того самца, который принимает участие в воспитании потомства (даже если это представитель другого вида, например, кенарь), и далее передают его своим потомкам. В результате можно получить несколько поколений снегирей, которые поют, как кенари. Существуют специфические влияния, связанные с гормональными сдвигами, которые прослеживаются между поколениями. Эти влияния возникают благодаря системе материнских гормонов, которые проникают через плацентарный барьер и оказывают воздействие на развивающийся плод. У человека социальные средовые факторы, которые приводят к возникновению стресса, бурно переживаемых эмоций, фрустрации, могут влиять на эндокринную систему матери и приводить к изменению пропорций определенных гормонов. Многие из этих гормонов могут проникать через плацентарный барьер в кровеносную систему плода и влиять на нервную и нейроэндокринную систему плода. В результате поведение новорожденного и его дальнейшее развитие могут отражать последствия тех социальных средовых влияний, которые испытывала мать в период беременности. Влияние пренатального стресса интенсивно исследуется на животных. Результаты этих исследований показывают, что ранний средовой стрессор может влиять не только на поведение взрослой самки, но оказывать воздействие и на дочерей. Кроме эмоционального стресса, также довольно интенсивно исследуется влияние недостаточного питания матери на развитие плода. В исследованиях Х. Бирча (Birch H.G., 1971, 1972, цит. по: Michel G.F., Moore C.L., 1995) было показано, что не только недостаточное питание матери во время беременности неблагоприятно сказывается на развитии ребенка, но и недостаточное питание самой матери в раннем детстве вызывает в ее организме необратимые сдвиги, которые в дальнейшем отрицательно отражаются на течении беременности и развитии ребенка, даже если будучи взрослой и при вынашивании ребенка мать питалась нормально. Описанный эффект был обнаружен Х. Бирчем в исследовании влияния социоэкономического статуса и расы на состояние младенцев при рождении (вес при рождении, смертность, заболеваемость). Ожидалось, что особенности младенцев будут коррелировать с социоэкономическим статусом матери, от которого зависит качество питания и медицинского обслуживания, получаемого матерью в период беременности. Вопреки ожиданию, состояние детей коррелировало не с социоэкономическим статусом матери, а с ее расой. Дети матерей негритянского происхождения, принадлежавших к среднему классу, демонстрировали более низкие показатели, чем дети белых матерей из того же класса. Состояние новорожденных у негритянских матерей среднего класса оказалось не лучше, чем состояние новорожденных у белых матерей, принадлежавших к низшему социальному классу. Результаты вызвали недоумение. Тогда Х. Бирч решил разделить негритянских матерей на две группы - тех, которые родились в условиях, характерных для жизни среднего класса, и тех, которые родились у родителей низшего класса и лишь затем волей обстоятельств стали принадлежать к среднему классу. Действительно, такое разделение привело к ожидаемым корреляциям состояния новорожденных с социоэкономическим статусом матерей. Таким образом, более существенным оказался социоэкономический статус матерей не в момент их беременности, а в момент их рождения. Это говорит о том, что пренатальный и ранний неонатальный опыт самой матери приводит к каким-то необратимым изменениям, которые затем сказываются в следующем поколении. Таким образом, есть основания полагать, что стрессогенные факторы и недостаточное питание в пренатальный и постнатальный период могут отрицательным образом сказываться на развитии ребенка, и если это девочка, то необратимые изменения, происходящие в организме в период раннего онтогенеза, могут неблагоприятным образом повлиять на ее потомство, даже если в последующие периоды жизни среда будет благополучной (см. также: Хрестомат. 9.2 и 9.3).
9.7. Принцип системности и диалектика взаимодействия генотипа и среды в развитии
9.7.1. Вариабельность в нервной системе
9.7.2. Эпигенез
9.7.3. Теория эпигенеза путем селективной стабилизации синапсов
9.7.4. Случайности развития
9.7.5. Историзм развития
Говоря о развитии, мы неоднократно упоминали о системности процесса развития и возможности его описания с помощью теории динамических систем. Особенно полезной теория систем оказывается при интерпретации процессов, сопровождающих морфогенез нервной системы и развитие различных форм поведения.
9.7.1. Вариабельность в нервной системе
Мы уже говорили,
что существует поразительное несоответствие
между относительно небольшим количеством
генов
в клеточном ядре и колоссальным
разнообразием клеточных элементов в
нервной системе. В гистологических
исследованиях мозга человека, наряду
со сходством основных принципов
клеточного строения (видоспецифических
особенностей), наблюдается значительная
межиндивидуальная вариабельность.
Можем
ли мы сравнить морфологическое строение
нервных клеток у близнецов? Чтобы сделать
это, необходимо в коре мозга каждого из
них отыскать идентичные по своему
происхождению клетки и сравнить их
фенотипы,
то есть детали строения - форму тела,
характер отростков, количество синапсов
и т.д. Понятно, что т акая
процедура практически неосуществима
для человека, но можно попытаться
проделать это для клонов других живых
организмов. Ж.-П. Шанже в своей книге
(Changeux
J.-P., 1997) приводит несколько
примеров, в которых сопоставляются
особенности строения нейронов-близнецов.
Например, удобно проделать это для
водяной блохи дафнии (Daphnia magna),
которая может размножаться партеногенетически
(без участия самца) и дает клоны организмов,
генетически идентичных материнскому.
Нервная система дафний примитивна,
каждый нейрон может быть
идентифицирован.
Оказалось,
что у клонов этих животных не наблюдается
вариабельности в количестве нервных
клеток. Например, глаз составлен ровно
из 176 сенсорных нейронов. Эти 176 нейронов
образуют контакты со 110 клетками
оптического ганглия. Здесь также нет
никаких вариаций. Каждый сенсорный
нейрон, взятый в отдельности, контактирует
с одними и теми же нейронами оптического
ганглия. Эта инвариантность, очевидно,
обеспечивается генетически. Однако
наряду с инвариантностью имеется и
определенная вариабельность: она
касается количества синапсов и формы
ветвления аксонов. Например, число
синапсов
одних и тех же нейронов у отдельных
особей может варьировать от 24 до 65.
Определенная вариабельность характерна
также для симметричных нейронов правой
и левой стороны тела, но она значительно
меньше вариабельности между особями
(рис. 9.11).
Подобные
наблюдения, сделанные для гигантских
нейронов рыб, также способных давать
изогенетические
клоны, дают сходные
результаты: межиндивидуальная
вариабельность у генетически идентичных
особей наблюдается лишь в мелких деталях
строения нейронов.
Можно
ли проделать подобные процедуры у
млекопитающих? К сожалению, на этот
вопрос придется пока дать отрицательный
ответ. Во-первых, возникают затруднения
с получением клонов животных в строгом
смысле этого слова. Можно, конечно,
воспользоваться инбредными
линиями и сопоставлять животных с почти
идентичными генотипами.
Однако возникает второе затруднение -
это идентификация одних и тех же нейронов
внутри определенных категорий. Число
нейронов здесь во много раз больше, чем
у простых организмов, и оно не является
строго фиксированным. Кроме того, нейроны
в процессе развития не занимают столь
регулярного и воспроизводимого в ряду
поколений положения, как у примитивных
дафний. Опыты с химерными мышами
показывают, что деление и миграция
нервных клеток у млекопитающих не
являются столь жестко детерминированными,
как у низших организмов, поэтому точное
расположение нервной клетки не является
строго фиксированным, и отыскать
"идентичные" нейроны в мозге
млекопитающих практически невозможно.
Отсюда понятно, что вариабельность
строения нервной системы млекопитающих
значительно возрастает и проявляется
не только в ветвлении отростков и
количестве синапсов, но и в числе и
местоположении нейронов. Поэтому тем
более не следует ожидать жесткой
воспроизводимости структуры у генетически
идентичных организмов. Вероятно,
возможности для фенотипической
вариабельности мозга у млекопитающих,
даже имеющих идентичные генотипы,
чрезвычайно велики.
Каким
же образом сложнейшая живая структура
- мозг человека - развивается из всего
одной клетки, имеющей весьма ограниченный
набор детерминант? Ответить на этот
вопрос невозможно, не привлекая теории
эпигенеза.
акая
процедура практически неосуществима
для человека, но можно попытаться
проделать это для клонов других живых
организмов. Ж.-П. Шанже в своей книге
(Changeux
J.-P., 1997) приводит несколько
примеров, в которых сопоставляются
особенности строения нейронов-близнецов.
Например, удобно проделать это для
водяной блохи дафнии (Daphnia magna),
которая может размножаться партеногенетически
(без участия самца) и дает клоны организмов,
генетически идентичных материнскому.
Нервная система дафний примитивна,
каждый нейрон может быть
идентифицирован.
Оказалось,
что у клонов этих животных не наблюдается
вариабельности в количестве нервных
клеток. Например, глаз составлен ровно
из 176 сенсорных нейронов. Эти 176 нейронов
образуют контакты со 110 клетками
оптического ганглия. Здесь также нет
никаких вариаций. Каждый сенсорный
нейрон, взятый в отдельности, контактирует
с одними и теми же нейронами оптического
ганглия. Эта инвариантность, очевидно,
обеспечивается генетически. Однако
наряду с инвариантностью имеется и
определенная вариабельность: она
касается количества синапсов и формы
ветвления аксонов. Например, число
синапсов
одних и тех же нейронов у отдельных
особей может варьировать от 24 до 65.
Определенная вариабельность характерна
также для симметричных нейронов правой
и левой стороны тела, но она значительно
меньше вариабельности между особями
(рис. 9.11).
Подобные
наблюдения, сделанные для гигантских
нейронов рыб, также способных давать
изогенетические
клоны, дают сходные
результаты: межиндивидуальная
вариабельность у генетически идентичных
особей наблюдается лишь в мелких деталях
строения нейронов.
Можно
ли проделать подобные процедуры у
млекопитающих? К сожалению, на этот
вопрос придется пока дать отрицательный
ответ. Во-первых, возникают затруднения
с получением клонов животных в строгом
смысле этого слова. Можно, конечно,
воспользоваться инбредными
линиями и сопоставлять животных с почти
идентичными генотипами.
Однако возникает второе затруднение -
это идентификация одних и тех же нейронов
внутри определенных категорий. Число
нейронов здесь во много раз больше, чем
у простых организмов, и оно не является
строго фиксированным. Кроме того, нейроны
в процессе развития не занимают столь
регулярного и воспроизводимого в ряду
поколений положения, как у примитивных
дафний. Опыты с химерными мышами
показывают, что деление и миграция
нервных клеток у млекопитающих не
являются столь жестко детерминированными,
как у низших организмов, поэтому точное
расположение нервной клетки не является
строго фиксированным, и отыскать
"идентичные" нейроны в мозге
млекопитающих практически невозможно.
Отсюда понятно, что вариабельность
строения нервной системы млекопитающих
значительно возрастает и проявляется
не только в ветвлении отростков и
количестве синапсов, но и в числе и
местоположении нейронов. Поэтому тем
более не следует ожидать жесткой
воспроизводимости структуры у генетически
идентичных организмов. Вероятно,
возможности для фенотипической
вариабельности мозга у млекопитающих,
даже имеющих идентичные генотипы,
чрезвычайно велики.
Каким
же образом сложнейшая живая структура
- мозг человека - развивается из всего
одной клетки, имеющей весьма ограниченный
набор детерминант? Ответить на этот
вопрос невозможно, не привлекая теории
эпигенеза.
9.7.2. Эпигенез
Развитие не является прямым следствием развертывания генетической программы. В предыдущих разделах этой главы мы пытались показать, насколько сложны и неоднозначны процессы взаимодействия различных субстанций внутри и вне клетки, насколько тесно они связаны со всеми событиями, которые происходят внутри и вне организма, - настолько, что трудно было бы представить, что в последовательности нуклеотидов могла бы быть записана вся программа развития. Эпигенез представляет развитие как системный динамический процесс, в котором гены могут играть роль своеобразных триггеров в процессах дифференцировки клеток, направляющих развитие клеток по тому или иному пути, но при этом существует множество негенетических факторов (клеточное окружение, поступление различных сигналов от других клеточных систем и из внешней среды, различные случайности развития и т.д.), которые модифицируют развитие. Жан-Пьер Шанже (Changeux J.-P., 1997) приводит пример с известной мутацией альбинизма, генетика которой хорошо изучена. Частота встречаемости этой мутации составляет один на 17 000 человек (рис. 9.12 а, б). Болезнь передается как рецессивный признак по простым менделевским законам. Мутация приводит к потере кожного пигмента меланина и, казалось бы, не должна затрагивать нервную систему. Однако тщательные анатомические исследования показывают, что у альбиносов имеются значительные изменения в строении латерального коленчатого тела и зрительного нерва. Здесь мы имеем дело с типичным плейотропным эффектом действия гена. Мутации, затрагивающие центральную нервную систему, чаще всего плейотропны. Альбинизм - одна из них. Как же возникает этот плейотропный эффект в случае альбинизма? Мы знаем, что альбиносы имеют красные глаза. Объясняется это тем, что сетчатка не содержит пигмента меланина, так же как и прочие клетки. Слой пигментных клеток в сетчатке появляется на ранних стадиях развития эмбриона, когда нейроны сетчатки еще продолжают делиться, а зрительный нерв еще не начал формироваться. В этот период эмбриональные нейроны сетчатки и пигментные клетки тесно соприкасаются. Вполне вероятно, что между этими слоями клеток происходит обмен сигналами. При этом, если пигмент присутствует, нейроны посылают свои аксоны в правильном направлении, и происходит нормальное формирование всего зрительного пути. Если пигмента нет, нервные волокна в зрительном нерве теряют упорядоченную организацию, беспорядочно перемешиваются и не дают того характерного перекреста, который существует в зрительном пути. В результате происходит реорганизация латерального коленчатого ядра и всего зрительного пути вплоть до коры мозга. Таким образом, в данной эмбриональной системе нарушение взаимодействия между двумя элементами в сети клеточных взаимодействий вызывает целый каскад изменений, затрагивающих другие элементы системы. В этом проявляется плейотропный эффект мутаций и вообще эффект единичного гена в развивающейся системе. Одним из важнейших этапов морфогенеза нервной системы является установление функциональных связей между клетками (как между самими нейронами, так и между нейронами и клетками тех органов и тканей, которые они иннервируют). Не нужно доказывать, что от характера нейронных сетей во многом будет зависеть и поведение индивида. Что же представляет собой эпигенез нервных связей? Какая роль здесь может быть отведена генам и какая - среде?
9.7.3. Теория эпигенеза путем селективной стабилизации синапсов
О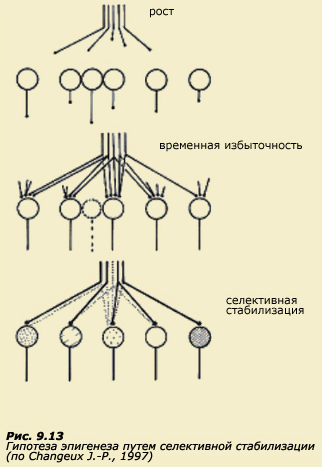 дной
из наиболее известных теорий эпигенеза
нервных связей является теория селективной
стабилизации синапсов Ж.-П. Шанже
(Changeux J.-Р., 1997). Согласно этой теории,
сохранение основных принципов организации
центральной нервной системы из поколения
в поколение, то есть сохранение
видоспецифических характеристик,
обеспечивается набором генов, которые
контролируют деление, миграцию и
дифференцацию нервных клеток; поведение
конуса роста; взаимное узнавание
клеточных категорий; образование широко
распространенных связей и начало
спонтанной активности. Фенотипическая
вариабельность, характерная даже для
генетически идентичных организмов,
особенно у высших форм млекопитающих,
включая человека, возникает в процессе
развития. Как только эмбриональные
нейроны заканчивают npоцесс деления,
начинается бурный и избыточный рост
нейритов (аксонов и дендритов) и их
активное ветвление. На этой критической
стадии наблюдается максимум избыточности
и разнообразия связей, устанавливаемых
в образующихся нейронных сетях. Эта
избыточность является временной. Вскоре
вступают в силу обратные процессы:
начинается гибель нейронов, значительная
часть нервных отростков исчезает,
исчезают и многие до того активные
синапсы (рис. 9.13).
Импульсы
начинают циркулировать в развивающихся
нервных сетях очень рано, с началом их
формирования. Сначала они возникают
спонтанно, но затем могут быть вызваны
взаимодействием растущего организма
со средой. В тот период, когда наблюдается
максимальная избыточность, эмбриональные
синапсы
могут существовать в трех состояниях:
лабильном, стабильном порождающемся.
Нервные импульсы передаются только
двумя первыми типами синапсов. Синапсы
могут переходить из одного состояния
в другое. Если лабильный синапс переходит
из лабильного состояния в стабильное,
то этот процесс называется стабилизацией.
Лабильный синапс может дегенерировать
(регрессия), а стабильный вновь стать
лабильным (лабилизация). По мере созревания
в процессе формирования функциональных
систем организма происходит постоянное
взаимодействие между всеми элементами
системы и внешней средой. В результате
функционально значимые связи остаются,
происходит стабилизация синапсов,
остальные (избыточные) связи
исчезают.
Процессы
регрессии, элиминации избыточности
являются типичными для всех уровней
развития нервной системы, в том числе
и поведенческого. Так, мы уже упоминали,
что с началом дыхательных движений
плода происходит упорядочение иннервации
межреберных мышц. Связи между полушариями
через мозолистое тело также подвержены
значительной регрессии, при этом
исключение какого-либо канала афферентации
(например, зрительного) ведет к сохранению
избыточности связей. Ж.-П. Шанже
предполагает, что процессы избирательной
элиминации на основе изначальной
избыточности лежат в основе обучения.
В качестве примеров на поведенческом
уровне он приводит процессы становления
речи. В доречевой период ребенок
продуцирует значительное количество
самых разнообразных звуков, из которых
только немногие представлены в речи
взрослого. В японском языке, например,
отсутствуют фонемы "г" и "1".
Взрослые японцы с трудом различают их,
однако 2-3-месячным младенцам их различение
дается настолько же легко, как и их
сверстникам из стран Запада.
Таким
образом, основы фенотипической
вариабельности закладываются изначально,
благодаря существованию механизмов,
создающих избыточность в нервной системе
(деление, миграция, дифференцировка,
избыточный рост и ветвление нейритов).
Последующие процессы регрессии и
селективной стабилизации зависят от
множества случайных событий развития,
от опыта конкретного индивида, который
не может быть абсолютно идентичным даже
для изогенных организмов. В результате
конечный фенотип будет определяться в
значительной степени процессами
взаимодействия элементов системы с
различными факторами, находящимися как
внутри, так и вне ее, то есть будет
зависеть от всей истории развития
индивида.
дной
из наиболее известных теорий эпигенеза
нервных связей является теория селективной
стабилизации синапсов Ж.-П. Шанже
(Changeux J.-Р., 1997). Согласно этой теории,
сохранение основных принципов организации
центральной нервной системы из поколения
в поколение, то есть сохранение
видоспецифических характеристик,
обеспечивается набором генов, которые
контролируют деление, миграцию и
дифференцацию нервных клеток; поведение
конуса роста; взаимное узнавание
клеточных категорий; образование широко
распространенных связей и начало
спонтанной активности. Фенотипическая
вариабельность, характерная даже для
генетически идентичных организмов,
особенно у высших форм млекопитающих,
включая человека, возникает в процессе
развития. Как только эмбриональные
нейроны заканчивают npоцесс деления,
начинается бурный и избыточный рост
нейритов (аксонов и дендритов) и их
активное ветвление. На этой критической
стадии наблюдается максимум избыточности
и разнообразия связей, устанавливаемых
в образующихся нейронных сетях. Эта
избыточность является временной. Вскоре
вступают в силу обратные процессы:
начинается гибель нейронов, значительная
часть нервных отростков исчезает,
исчезают и многие до того активные
синапсы (рис. 9.13).
Импульсы
начинают циркулировать в развивающихся
нервных сетях очень рано, с началом их
формирования. Сначала они возникают
спонтанно, но затем могут быть вызваны
взаимодействием растущего организма
со средой. В тот период, когда наблюдается
максимальная избыточность, эмбриональные
синапсы
могут существовать в трех состояниях:
лабильном, стабильном порождающемся.
Нервные импульсы передаются только
двумя первыми типами синапсов. Синапсы
могут переходить из одного состояния
в другое. Если лабильный синапс переходит
из лабильного состояния в стабильное,
то этот процесс называется стабилизацией.
Лабильный синапс может дегенерировать
(регрессия), а стабильный вновь стать
лабильным (лабилизация). По мере созревания
в процессе формирования функциональных
систем организма происходит постоянное
взаимодействие между всеми элементами
системы и внешней средой. В результате
функционально значимые связи остаются,
происходит стабилизация синапсов,
остальные (избыточные) связи
исчезают.
Процессы
регрессии, элиминации избыточности
являются типичными для всех уровней
развития нервной системы, в том числе
и поведенческого. Так, мы уже упоминали,
что с началом дыхательных движений
плода происходит упорядочение иннервации
межреберных мышц. Связи между полушариями
через мозолистое тело также подвержены
значительной регрессии, при этом
исключение какого-либо канала афферентации
(например, зрительного) ведет к сохранению
избыточности связей. Ж.-П. Шанже
предполагает, что процессы избирательной
элиминации на основе изначальной
избыточности лежат в основе обучения.
В качестве примеров на поведенческом
уровне он приводит процессы становления
речи. В доречевой период ребенок
продуцирует значительное количество
самых разнообразных звуков, из которых
только немногие представлены в речи
взрослого. В японском языке, например,
отсутствуют фонемы "г" и "1".
Взрослые японцы с трудом различают их,
однако 2-3-месячным младенцам их различение
дается настолько же легко, как и их
сверстникам из стран Запада.
Таким
образом, основы фенотипической
вариабельности закладываются изначально,
благодаря существованию механизмов,
создающих избыточность в нервной системе
(деление, миграция, дифференцировка,
избыточный рост и ветвление нейритов).
Последующие процессы регрессии и
селективной стабилизации зависят от
множества случайных событий развития,
от опыта конкретного индивида, который
не может быть абсолютно идентичным даже
для изогенных организмов. В результате
конечный фенотип будет определяться в
значительной степени процессами
взаимодействия элементов системы с
различными факторами, находящимися как
внутри, так и вне ее, то есть будет
зависеть от всей истории развития
индивида.
9.7.4. Случайности развития
Вариабельность феноменов развития зависит от многих причин. Наследственность имеет тенденцию уменьшать вариабельность развития, тогда как условия, не связанные с наследственностью, имеют тенденцию ее повышать. Некоторые исследователи развития выделяют четыре типа случайных факторов, которые влияют на вариабельность развития:
случайности в подборе родительских пар, гены которых слагают генотип индивида;
случайности эпигенетических (то есть внешних по отношению к генотипу) процессов в пределах индивидуального онтогенеза;
случайности материнской среды, в которой развивается индивид;
случайности нематеринской среды, в которой развивается индивид.
Хотя это и случайные события, однако, все они имеют элемент наследственности. Генотип наследуется от родителей, и у потомка с родителями имеются общие гены, которые влияют на ход индивидуального развития. Эпигенетические процессы внутри организма представляют собой влияния других клеток или их продуктов на активность генотипа данной клетки. Поскольку все клетки организма имеют один и тот же генотип, естественно, что эпигенетические влияния связаны с наследственностью. Однако эпигенетические процессы являются стохастическими, открытыми влиянию факторов среды организма и, следовательно, любым историческим случайностям. Материнская среда млекопитающих является очень важным элементом внешней среды. Матери обеспечивают внутриутробную и постнатальную (уход за младенцем и воспитание) среду ребенка. Понятно, что на эти условия действует генотип матери. Частично же гены матери являются общими с потомком, поэтому материнская среда может наследоваться. Материнская среда также чувствительна к историческим случайностям. Нематеринские средовые эффекты также влияют на вариабельность развития. Сюда входят факторы, которые выбираются самим индивидом или формируются окружающими людьми, в том числе родственниками, с которыми у него имеются общие гены. Поэтому и эти средовые эффекты в какой-то мере также находятся не только под влиянием случайных средовых событий, но и под влиянием генов, и также наследуются (генотип-средовая ковариация). Таким образом, в соответствии с приведенной классификацией во всех описанных элементах внешней по отношению к данному индивиду среды имеются механизмы для наследования как генетического, так и негенетического (различные традиции и т.п.). Естественно, на развитие действуют и ненаследуемые факторы. Это те особенности среды, которые не связаны с изменениями, вызываемыми самим развивающимся индивидом или его родственным окружением. Они могут быть как случайными, так и закономерными. К закономерным можно отнести циклические изменения (смена дня и ночи, смена времен года и т.п.), повсеместные воздействия (гравитация) или предсказуемые факторы (температура, давление). Ненаследуемые факторы присутствуют также в материнской и другой социальной среде (качество питания матери, уровень стресса матери, число и пол сиблингов и др.). Случайно или систематически изменяющиеся средовые события способствуют вариативности развития. Все внешние по отношению к генам события, которые имеют место в процессе онтогенеза, в совокупности с генетическими факторами создают тот фон, на котором протекает развитие. Благодаря воздействию огромного разнообразия закономерных и случайных событий в онтогенезе, развивающиеся системы могут организовываться и реорганизовываться. Гены делают развитие возможным, но и другие компоненты, влияющие на развитие системы, являются не менее важными участниками процесса развития. В начале изложения, определяя понятие фенотипа, мы подчеркивали, что фенотип является результатом взаимодействия генотипа и среды, однако в свете того, что было сказано о процессе индивидуального развития, мы должны внести некоторое уточнение в эту формулировку и, наряду с факторами среды, упомянуть о случайностях развития, которые не могут быть сведены к чисто средовым влияниям. Если бы мы попытались графически изобразить зависимость фенотипа от различных факторов (аналогично зависимости фенотипа от среды, когда мы рассматривали норму реакции - см. рис. 9.1), то нам понадобилось бы по крайней мере четырехмерное пространство, в котором, помимо осей для генотипа и среды, обязательно должна была бы присутствовать и ось для случайностей развития (см. также: Хрестомат. 9.4).
9.7.5. Историзм развития
Как подчеркивает Р. Левонтин (1993), нельзя раз и навсегда охарактеризовать индивидуальные особенности человека, описав все характерные черты его фенотипа (биохимические, морфологические, физиологические, поведенческие). Нет ни одной системы организма, которая не находилась бы на протяжении жизни в процессе постоянного развития и обновления. Это относится ко всем уровням функционирования, начиная с генного и кончая поведенческим. Каждый индивид в любой конкретный момент его жизни представляет собой результат всех событий, происходивших в истории его развития. Лишь проследив и описав историю всех изменений, произошедших в организме человека и в его поведении в течение всей его жизни, можно получить достаточно полное описание данного индивида. Все изменения, происходящие в процессе развития, зависят от текущего состояния организма и условий среды, в которых он находится. Невозможно предсказать, каким будет организм в определенный момент времени, зная лишь одну переменную (скажем, параметры среды), необходимо иметь сведения и о второй переменной (состояние систем организма). Отсюда важное следствие - одно и то же средовое воздействие может вызвать разные последствия в зависимости от стадии онтогенеза, на которой оно было осуществлено. Известно, например, что алкоголь и некоторые лекарственные препараты, действующие на человеческий плод на ранних этапах развития, могут привести к тяжелым и необратимым последствиям, тогда как для взрослого организма они относительно безопасны. Вторым важным следствием историзма развития является то, что реакция на воздействие конкретной среды в каждый конкретный момент времени определяется не просто текущим состоянием индивида, но зависит от истории всех средовых воздействий, которым индивид подвергался на протяжении предыдущего периода. Каждое новое средовое воздействие "ложится" не на чистый генотип, а на результат предыдущих взаимодействий, которые уже "записаны" во всех системах организма. Развитие зависит не просто от условий среды, а от истории смены этих условий. В этом суть детерминированности сегодняшнего состояния организма и его возможного будущего прошлым опытом. Информация о прошлом опыте может храниться в различных системах "памяти" организма (на уровне генов, на уровне иммунной системы, в памяти нервной системы). С иммунологической памятью, в частности, связаны различные аллергические реакции организма. Точно так же любые психологические характеристики, например способности, представляют собой черты поведения, которые формируются на протяжении всей жизни и зависят от истории смены всех обстоятельств развития, информация о которых записывается в памяти нервной системы.
Выводы Исследования развития в психогенетике ведутся на популяционном уровне; получаемые в результате количественные соотношения генетических и средовых компонент изменчивости неприложимы к развитию конкретного фенотипа. Необходимо помнить, что взаимовлияния генотипа и среды в индивидуальном развитии неразделимы. Формирование фенотипа в развитии происходит при непрерывном взаимодействии генотипа и среды. Факторы внешней среды (физические, социальные) могут влиять на генотип через факторы внутренней среды организма (различные биохимические субстанции внутри клетки). Основным механизмом взаимодействия генотипа и среды на уровне клетки является регуляция экспрессии гена, проявляющейся в разной активности синтеза специфического белка. Большая часть процессов регуляции происходит на уровне транскрипции, то есть касается процессов считывания генетической информации, необходимой для синтеза белка. Среди всех органов тела мозг занимает первое место по количеству активных генов. По некоторым оценкам почти каждый второй ген в геноме человека связан с обеспечением функций нервной системы. Ранний опыт имеет значительные возможности влиять на работу генетического аппарата. Особая роль здесь принадлежит так называемым ранним генам, которые способны к быстрой, но преходящей экспрессии в ответ на сигналы из внешней среды. По всей видимости, ранние гены играют значительную роль в процессах обучения. Значительные возможности регуляции экспрессии генов связаны также с действием различных гормонов. Развитие нервной системы и в конечном счете поведения представляет собой динамический иерархически организованный системный процесс, в котором в равной степени важны генетические и средовые факторы. Немаловажную роль играют также различные случайности развития, которые не могут быть сведены к чисто средовым. Развитие представляет собой эпигенетический процесс, приводящий к формированию значительной межиндивидуальной вариабельности даже у изогенных организмов. Основным принципом морфогенеза нервной системы является возникновение максимальной избыточности клеточных элементов и их связей на ранних этапах развития, с последующей элиминацией функционально нестабильных элементов в процессе реципрокного взаимодействия между всеми уровнями развивающейся системы, включая взаимодействия внутри клетки, между клетками и тканями, между организмом и средой. Процесс формирования фенотипа в развитии носит непрерывный диалектический и исторический характер. На любом этапе онтогенеза характер реакции организма на воздействие среды определяется как генотипом, так и историей всех обстоятельств развития.
Выводы
Исследования развития в психогенетике ведутся на популяционном уровне; получаемые в результате количественные соотношения генетических и средовых компонент изменчивости неприложимы к развитию конкретного фенотипа. Необходимо помнить, что взаимовлияния генотипа и среды в индивидуальном развитии неразделимы.
Формирование фенотипа в развитии происходит при непрерывном взаимодействии генотипа и среды. Факторы внешней среды (физические, социальные) могут влиять на генотип через факторы внутренней среды организма (различные биохимические субстанции внутри клетки).
Основным механизмом взаимодействия генотипа и среды на уровне клетки является регуляция экспрессии гена, проявляющейся в разной активности синтеза специфического белка. Большая часть процессов регуляции происходит на уровне транскрипции, то есть касается процессов считывания генетической информации, необходимой для синтеза белка.
Среди всех органов тела мозг занимает первое место по количеству активных генов. По некоторым оценкам почти каждый второй ген в геноме человека связан с обеспечением функций нервной системы.
Ранний опыт имеет значительные возможности влиять на работу генетического аппарата. Особая роль здесь принадлежит так называемым ранним генам, которые способны к быстрой, но преходящей экспрессии в ответ на сигналы из внешней среды. По всей видимости, ранние гены играют значительную роль в процессах обучения. Значительные возможности регуляции экспрессии генов связаны также с действием различных гормонов.
Развитие нервной системы и, в конечном счете, поведения представляет собой динамический иерархически организованный системный процесс, в котором в равной степени важны генетические и средовые факторы. Немаловажную роль играют также различные случайности развития, которые не могут быть сведены к чисто средовым.
Развитие представляет собой эпигенетический процесс, приводящий к формированию значительной межиндивидуальной вариабельности даже у изогенных организмов. Основным принципом морфогенеза нервной системы является возникновение максимальной избыточности клеточных элементов и их связей на ранних этапах развития, с последующей элиминацией функционально нестабильных элементов в процессе реципрокного взаимодействия между всеми уровнями развивающейся системы, включая взаимодействия внутри клетки, между клетками и тканями, между организмом и средой.
Процесс формирования фенотипа в развитии носит непрерывный диалектический и исторический характер. На любом этапе онтогенеза характер реакции организма на воздействие среды определяется как генотипом, так и историей всех обстоятельств развития.
Словарь терминов
Индивидуальное развитие
Норма реакции
Генотип
Среда
Фенотип
Ранний опыт
Фенотип клетки
Белок
Ген
Кодон
Экзон
Интрон
Транскрипция
Трансляция
РНК
Экспрессия гена
Сплайсинг
Рибосома
Ранние гены
Гормоны
G-белки
Эмбриогенез
Морфогенез
Функциональная система
Реципрокное взаимодействие
Рождение нейронов
Миграция нейронов
Дифференциация нейронов
Гибель нейронов
Трофические факторы
Фактор роста нервов (ФРН)
Нейрит
Аксон
Дендрит
Пластичность нервной системы
Гаметогенез
Эмбриогенез
Морфогенез
Органогенез
Пренатальный
Неонатальный
Постнатальный
Эмбрион
Плод
Новорожденный
Мутация
Кроссинговер
Генетический импринтинг
Материнский эффект
Хэндлинг
Межпоколенные эффекты
Вариабельность развития
Случайности развития
Преформация
Эпигенез
Эпигенетические факторы
Системность развития
Критический период
Сензитивный период
Кризисный возраст
Вопросы для самопроверки
Можно ли по данным психогенетических исследований прогнозировать фенотип конкретного индивида?
Можно ли судить о генетических и средовых причинах индивидуального развития, имея данные о соотношении наследственного и средового компонента в вариативности данного признака?
Поясните на графическом примере, как высокая наследуемость может сочетаться с влияниями среды на ход индивидуального развития.
Имеются ли возможности для влияний среды на формирование фенотипа при 100%-ной наследуемости?
Почему норма реакции не должна определяться через понятие предела?
Как вы думаете, верно ли утверждение, что пределы фенотипа непознаваемы?
Как вы представляете себе процесс взаимодействия генотипа и среды в индивидуальном развитии?
Что входит в понятие фенотипа клетки?
Что бы вы включили в фенотип нервной клетки?
Почему основу клеточного фенотипа составляют белки?
Какие особенности строения белка определяют его специфику?
Какую роль играет ДНК в функционировании клетки?
Почему формула "один ген - один фермент" не может считаться верной?
Что такое процессы транскрипции и трансляции и где они происходят?
Какие агенты участвуют в процессах транскрипции и трансляции?
Что такое экспрессия гена?
На каких ступенях экспрессии гена возможна ее регуляция?
Что может быть результатом регуляции экспрессии гена?
Какими молекулярно-генетическими механизмами можно объяснить огромное разнообразие клеток и клеточных продуктов в нервной системе?
Назовите основные механизмы регуляции экспрессии генов.
Что такое ранние гены и какую роль они играют в развитии?
Какую роль в генетической регуляции играют гормоны и почему?
Опишите процессы клеточной регуляции, идущие при участии G-белков.
Как вы представляете себе процесс взаимодействия генотипа и среды на разных уровнях: клеточном, организменном, внеорганизменном?
Почему особенности поведения во многом определяются этапами раннего развития нервной системы?
Что представляет собой функциональная система?
Как вы представляете себе реципрокные взаимодействия между растущей нервной системой и прочими системами организма?
Какие события связаны с развитием отдельной клетки в нервной системе?
Какими процессами регулируется общее количество нервных клеток в организме?
Что известно о процессах миграции и дифференциации нервных клеток?
Какую роль в развитии нервной системы играют процессы взаимодействия с другими клетками?
Как растущие нейроны устанавливают функциональные связи?
Что такое трофические факторы?
Как вы представляете себе работу генов в процессе роста и дифференциации нервной ткани?
Какую роль в процессе развития нервной системы играет естественная гибель клеток?
Может ли внешняя среда регулировать процессы морфогенеза нервных клеток? Как вы это представляете?
Почему развивающийся организм относится к динамическим системам?
Почему развитие можно считать непрерывным процессом?
Какие типы движений характерны для плода человека и какова их возможная роль?
Какие сенсорные системы начинают функционировать еще до рождения?
Как факторы среды могут влиять на развитие поведения во внутриутробном периоде?
Что известно о влиянии раннего слухового опыта плода на поведение новорожденного?
Как ранний вестибулярный опыт плода может влиять на закладку функциональной асимметрии мозга?
Благодаря чему самый ранний опыт может существенно влиять на последующее развитие поведения?
Как вы думаете, может ли поведение младенца объясняться генетическими программами?
На какие процессы развития может влиять физиологическое состояние родителей?
Какие генетические процессы чувствительны к физиологическому состоянию родителей?
Что представляет собой явление генетического импринтинга?
Что такое материнский эффект и из каких элементов он складывается?
Какие эксперименты позволяют развести отдельные составляющие материнского эффекта?
Как вы представляете себе пренатальный и постнатальный материнский эффект у человека?
Что такое реципрокные взаимодействия матери и ребенка и на какие системы они влияют?
Какие факторы материнской среды могут иметь отдаленные последствия?
Влияние каких материнских факторов наиболее изучено и что вы об этом знаете? Какие факторы повышают и какие понижают вариабельность развития?
Какие типы случайных факторов влияют на вариабельность развития?
Почему некоторые случайные события в развитии несут элемент наследственности? Как вы это себе представляете?
Какие закономерные и случайные ненаследуемые факторы развития вы можете назвать?
Как вы представляете себе системность развития?
Что понимается под исторической природой развития?
Как можно представить себе модель развития в четырехмерном пространстве?
Какую роль в развитии играет прошлый опыт организма?
Что вы знаете о сензитивных и критических периодах развития? На какие периоды онтогенеза они приходятся?
Раздел VI. Психогенетические исследования нормальной вариативности
В психогенетике (генетике поведения человека) можно выделить два крупных направления исследований. Одно из них разрабатывается в основном психологами и связано с изучением наследственных и средовых причин изменчивости нормальных психологических и психофизиологических признаков (интеллект, темперамент, личность, сенсорные и двигательные функции, психофизиологические характеристики). Второе направление связано с изучением наследственных и средовых причин психических заболеваний и различных отклонений в развитии психики, а также различных форм девиантного поведения. В разработке последнего направления принимают участие как медики, так и психологи. Сложилось так, что основное внимание исследователей генетики поведения человека было сосредоточено на изучении сложных мультифакториальных признаков и заболеваний. В исследованиях нормальной вариативности преобладали исследования интеллекта и когнитивных характеристик (около 80% всех работ), в психиатрии главное внимание было направлено на такие заболевания, как шизофрения, аффективные расстройства, умственная отсталость. В последние годы появился интерес к изучению нарушений развития (нарушение способности к обучению, ранний детский аутизм, синдром гиперактивности и дефицита внимания) и различных форм отклоняющегося поведения (агрессивное поведение, употребление алкоголя и наркотических веществ, гомосексуализм и т.п.). Как ни странно, очень мало исследованы более простые функции, которые не столь далеко отстоят от генного уровня и, по-видимому, должны находиться в большей зависимости от наследственных влияний: морфология и физиология мозга, различные виды сенсорной чувствительности, восприятия и двигательных функций. В данном разделе мы постараемся познакомить читателя с основными достижениями психогенетики в изучении вариативности нормальных психических и психофизиологических характеристик человека и начнем с более простых признаков, среди которых есть и наследуемые по Менделю.
Тема 10. Элементарные психические функции. Психофизиологические и двигательные характеристики
10.1. Сенсорное восприятие
10.2. Морфология и физиология мозга
10.3. Двигательные характеристики
10.1. Сенсорное восприятие
Одним из наиболее известных видов наследственного полиморфизма у человека является разная чувствительность к вкусу фенилтиомочевины (ФТМ). Фенилтиомочевина представляет собой бесцветное кристаллическое вещество. Некоторые люди ощущают вкус этого вещества даже при малых концентрациях: оно кажется им интенсивно горьким. Другие люди считают его безвкусным. Существуют вещества, похожие по химическому составу, вкус которых воспринимается таким же образом. По-видимому, ощущение вкуса зависит от способности вкусовых рецепторов воспринимать группировку = N - C = S. В 1931 г. было обнаружено, что способность ощущать вкус ФТМ является наследственным признаком. Оказалось, что признак наследуется по Менделю. Рецессивные гомозиготы (tt) не ощущают вкус ФТМ, тогда как гетерозиготы (Tt) и гомозиготы с доминантными аллелями (ТТ) считают ФТМ горьким веществом. Этот тип вкусового полиморфизма свойствен не только человеку, но и большинству приматов. Нечувствительные к вкусу ФТМ люди чаще страдают заболеваниями щитовидной железы, среди них чаще встречаются индивиды, которым не нравится вкус алкогольных напитков. Примерно 25-30% людей имеют генотип tt. Хотя установлен рецессивный тип наследования нечувствительности к ФТМ, однако существуют и другие возможные объяснения, включая неполное доминирование и множественный аллелизм. Райф (Rife, 1938; цит. по: Эрман Л. и Парсонс П., 1984), например, обнаружил почти 4% дискордантность МЗ близнецов по чувствительности к вкусу ФТМ. Хотя наследственный характер чувствительности к ФТМ был установлен достаточно давно, соответствующий ген удалось выявить лишь в 1999 г. (Reed D.L. et al., 1999) в результате анализа сцепления в 98 семьях с использованием 356 маркеров, расположенных с интервалами порядка 10 сМ (сантиморганид). Главным кандидатом является ген, расположенный на коротком плече 5 хромосомы. Еще один возможный ген-кандидат может быть связан с 7-й хромосомой. Исследовались также генетические аспекты других вкусовых ощущений. Было обнаружено три вкусовых локуса для разных веществ, причем в одном из них объединялись такие вещества, как хинин, сахароза и хлористый натрий. Оказалось также, что чем ниже был порог ощущения горьких веществ, тем более разборчивым оказывался данный индивид по отношению к различным видам пищи. Пороги вкусовой чувствительности в семьях значительно варьируют. Наследственные особенности обоняния, кинестетической и тактильной чувствительности у человека остаются практически не изученными. Имеются лишь некоторые указания на существование наследственных форм нечувствительности к запахам ряда веществ. Генетика вариативности нормальной слуховой чувствительности также почти не исследована, хотя работ по изучению наследственной передачи глухоты и тугоухости достаточно. Что касается нормы, то здесь основное внимание было привлечено к наследованию абсолютного музыкального слуха. Абсолютный музыкальный слух определяется как способность безошибочно определять высоту звука или нескольких звуков при прослушивании и точно называть ноту. Люди, обладающие абсолютным слухом, часто имеют и феноменальную музыкальную память. Помимо этого для них характерны высокие математические способности и хорошая память вообще. У обладателей абсолютного слуха иногда встречаются и другие виды сенсорной одаренности (в области вкусовых ощущений, например). Некоторые из них способны к синестезиям (цвето-музыкальным ассоциациям). Известно, что такой способностью обладали такие композиторы, как Скрябин, Сибелиус, Чюрленис. Абсолютный слух чаще встречается у женщин. Имеются указания на то, что абсолютный слух коррелирует с нарушениями способности к обучению в школе (learning disability). Абсолютный слух сопутствует также некоторым наследственным неврологическим заболеваниям (синдром Виллиса, аутизм). У музыкантов с абсолютным слухом (правшей) была выявлена более значительная межполушарная асимметрия в височной области коры головного мозга: размеры левой planum temporale - височной пластинки - оказались значительно больше, чем правой. Современные компьютерные методы исследования мозга показывают, что у обладателей абсолютного слуха имеются и другие особенности мозговой организации, отличающие их от остальной популяции. В проведенном в 1998 г. исследовании (Baharlow и др.), включающем 600 профессиональных музыкантов, была сделана попытка проверить, не влияет ли ранний опыт на развитие абсолютного слуха. Оказалось, что раннее начало музыкальных занятий (в 4 года и раньше), способствует развитию абсолютного музыкального слуха. По-видимому, помимо наследственной предрасположенности, на развитие абсолютного слуха влияет и научение. В 2000 г. Baharlou и др. было проведено формализованное тестирование восприятия высоты звука на выборке испытуемых и отобраны индивиды с особенно высокими результатами. Тестирование сибсов этих испытуемых показало, что исключительно высокие способности к восприятию высоты звука имеют тенденцию к накоплению в семьях. Как считают авторы, при достаточном количестве родственников, можно будет картировать локус, отвечающий за абсолютный слух. Генетика зрительной чувствительности и зрительного восприятия также изучены чрезвычайно слабо. Наиболее известны наследственные варианты нарушений цветового зрения. Неспособность различать красный и зеленый цвета у некоторых людей была отмечена еще в XVIII в. Это нарушение известно под названием "дальтонизм". Большинство людей различают цвета, полученные в результате смешения красного, зеленого и синего частей спектра. Их называют трихроматами. У некоторых людей в сетчатке отсутствуют элементы для восприятия одного из цветов. Такое нарушение носит название дихромазии. Слепота на красный цвет называется протанопия, а слепота на зеленый - дейтеранопия. Оба нарушения контролируются двумя сцепленными с полом рецессивными аллелями, поэтому их частота гораздо выше у мужчин (в среднем 8 %), чем у женщин (0,64 %). Встречается также нарушение различения цветов в сине-зеленой области, однако сцепления с полом здесь не наблюдается. Признак, по-видимому, контролируется аутосомным геном. Частота встречаемости мужчин с нарушениями цветового зрения варьирует в популяциях (примерно от 2% до 12%), причем она выше в индустриально развитых странах. Цветовая слепота может развиваться и у людей, не обладающих соответствующим генотипом. Показано, что на цветовое зрение могут влиять некоторые психические особенности человека, в частности, плохая психическая приспособляемость и плохая приспособляемость к особенностям питания (Dunlop, 1943; цит. по: Эрман Л., Парсонс П., 1984). В этом случае возникают фенокопии цветовой слепоты. Фенокопиями называются модификации фенотипа, развивающиеся под действием среды, и имитирующие наследственные варианты. Как ни странно, среди учащихся художественных школ довольно часто встречаются дихроматы. Если студент уверен в себе и не знает о своем дефекте, его недостаток мало сказывается на его успехах. Часто такие учащиеся работают в оригинальной цветовой гамме. В психогенетике исследовались также некоторые особенности зрительного восприятия, включая отдельные иллюзии, размер остаточного изображения, критическую частоту мельканий, время опознания изображения. Часто методики применяются в исследованиях, не направленных непосредственно на изучение наследственных причин вариативности зрительного восприятия как такового, а в рамках других задач. Например, некоторые скоростные особенности зрительного восприятия рассматриваются как корреляты интеллектуальных способностей и изучаются в рамках исследований наследуемости интеллекта. Ряд методик используется в психогенетических исследованиях свойств нервной системы (например, критическая частота мельканий). Хотя применяемые экспериментальные подходы и количество испытуемых значительно варьируют, в целом результаты демонстрируют наличие довольно существенного вклада наследственности в особенности зрительного восприятия. В качестве иллюстрации приведем таблицу внутриклассовых корреляций МЗ и ДЗ близнецов и значений наследуемости для некоторых тестов зрительного восприятия (табл. 10.1).
Таблица 10.1
Внутриклассовые корреляции и значения h2-статистик для некоторых тестов на особенности восприятия по данным о монозиготных и дизиготных близнецах (по Эрман Л., Парсонс П., 1984)
Тест |
|
rМЗ |
rДЗ |
h2 |
Размер остаточного изображения |
(1) (2) (3) (4) |
0,71 0,68 0,98 0,75 |
0,08 0,00 0,22 0,23 |
0,68 0,68 0,97 0,67 |
Внимательность |
(1) (2) (3) |
0,50 0,66 0,67 |
0,10 0,15 0,05 |
0,44 0,60 0,65 |
Критическая частота мельканий |
|
0,71 |
0,21 |
0,63 |
Иллюзия Мюллера-Лайера |
(1) (2) (3) (4) |
0,53 0,55 0,51 0,57 |
0,39 0,05 0,37 0,28 |
0,22 0,52 0,22 0,40 |
Автокинетический феномен |
|
0,72 |
0,21 |
0,64 |
Несмотря на весьма скудные сведения о наследственных и средовых причинах изменчивости сенсорных характеристик человека, можно все же полагать, что люди по-разному воспринимают мир, и в основе этих различий лежат не только особенности среды, в которых происходит развитие. Вероятно, существуют определенные наследственные варианты сенсорной чувствительности и сенсорного восприятия, которые вместе с разнообразием сред формируют наблюдаемые индивидуальные различия. Каждый человек, по-видимому, воспринимает одни и те же сигналы окружающей среды по-своему. Соответственно и реакции разных людей на одни и те же явления и события могут отличаться в силу различий, существующих на уровне элементарных ощущений и восприятия.
10.2. Морфология и физиология мозга
10.2.1. Генетические исследования морфологии мозга человека
10.2.2. Генетические исследования биоэлектрической активности мозга человека
10.2.1. Генетические исследования морфологии мозга человека
Особенности строения человеческого мозга постоянно привлекают внимание ученых. Многочисленные исследования post mortem демонстрируют поразительную вариабельность морфологии мозга. Значительные межиндивидуальные различия наблюдаются в характере борозд и извилин, распределении белого и серого вещества, в строении подкорковых структур и цитоархитектонике мозга (Блинков С.М., Глезер И.И., 1964). Отмечаемое своеобразие в строении мозга каждого индивида является результатом сложнейших взаимодействий наследственных и средовых факторов, а также многочисленных случайных событий, сопутствующих развитию нервной системы в онтогенезе. Об этом мы будем более подробно говорить в теме 12. Фенотипические различия в объеме мозга, по-видимому, в значительной степени связаны с генотипической вариативностью. Об этом свидетельствует исследование, выполненное недавно в Нидерландах (Baare W.F.C. et al., 2001). Исследовались близнецы и их сибсы - 54 пары МЗ и 58 пар ДЗ близнецов, а также их 34 сибса. Был применен метод магнитно-резонансного сканирования мозга (MRI) с высокой разрешающей способностью. Это первое исследование такого рода, выполненное на должном методическом уровне. Ранее проводились либо исследования посмертного материала на очень небольших выборках, либо применялись методы компьютерной томографии и MRI in vivo лишь на группах МЗ близнецов (без контрольной группы ДЗ), что не позволяло разделить наследственные и средовые компоненты наблюдаемого сходства. Для количественной оценки вклада наследственных и средовых компонентов в вариативность таких показателей, как общий объем мозга, объем серого и белого вещества и некоторых других характеристик, в нидерландском исследовании были применены методы структурного моделирования (см. тему 5). Результаты показали, что вклад аддитивного генетического компонента в вариативность характеристик объема и размеров мозга очень высок и составляет от 82 до 90%. Остальная вариативность возникает за счет индивидуальной среды. Роль общей среды оказалась незначимой. Единственным показателем, который описывался чисто средовой моделью, оказался объем латерального желудочка мозга (58% составил вклад общей среды и 42% - вклад индивидуальной среды). Авторы считают, что параметры объема мозга можно рассматривать как промежуточный фенотип между генами и поведением в психогенетических исследованиях. Подобие и различие морфологического строения мозга у близнецов и их сибсов приведено на рис. 10.1. На рис. 10.2 приводятся соответствующие диаграммы путей (см. тему 5).
Представленное выше исследование выполнено благодаря развитию новых неинвазивных технологий, применяемых в настоящее время для изучения мозга. Можно ожидать, что это направление получит дальнейшее развитие, и в скором времени мы сможем получить новые данные, касающиеся генетики строения мозга человека. По-видимому, количественные параметры объема мозга в значительной степени варьируют за счет генетических причин.
10.2.2. Генетические исследования биоэлектрической активности мозга человека
Помимо морфологического разнообразия наблюдается значительная вариативность и в функционировании мозга человека, что проявляется в специфике биоэлектрической активности. Наиболее распространенными методами исследования биоэлектрической активности мозга являются регистрация электроэнцефалограммы (ЭЭГ) и связанных с событиями потенциалов мозга, или вызванных потенциалов (слуховых, зрительных, соматосенсорных и связанных с движениями). Как ЭЭГ, так и вызванные потенциалы (ВП) характеризуются своеобразным рисунком, отличающимся у разных индивидов, но, в то же время, сохраняющим индивидуальную специфику, что выявляется при повторной регистрации этих видов активности у одних и тех же людей. Такие особенности биоэлектрических паттернов мозга позволили предположить, что в индивидуальных особенностях функционирования мозга проявляются наследственные черты. Методы регистрации ЭЭГ и ВП начали применяться достаточно давно - еще в первой половине ХХ в. В 1929 г. Г. Бергер зарегистрировал волновую активность мозга у человека. Достаточно регулярные волны с частотой около 10 колебаний в секунду, доминирующие в ЭЭГ большинства людей в состоянии покоя, были названы им "альфа-ритмом" (ритм Бергера). В 1939 г. Дэвис, регистрируя ЭЭГ, обнаружил большую негативную волну, появляющуюся через 100-200 мс после подачи звукового стимула. С тех пор методы регистрации ЭЭГ и ВП мозга стали неотъемлемой частью нейрофизиологических исследований. (Более подробно с этими методами можно познакомиться в руководствах по нейро- и психофизиологии.) Примерно в те же годы возник вопрос о происхождении устойчивых индивидуальных различий в биоэлектрической активности мозга. Можно было предположить, что различные индивидуальные варианты паттернов ЭЭГ и ВП имеют наследственную природу. Чтобы проверить это предположение, исследователи обратились к методу близнецов. Исследования электроэнцефалограммы в психогенетике. Электроэнцефалограмма (ЭЭГ) есть результат регистрации разности электрических потенциалов между различными точками кожного покрова головы. Она представляет собой сумму ритмических колебаний различной частоты. В зависимости от частоты выделяют различные ритмические составляющие ЭЭГ: дельта-волны с частотой до 4 Гц, тета-волны с частотой 4-8 Гц, альфа-волны с частотой 8-13 Гц, бета-волны с частотой 15-50 Гц. В состоянии покоя в темноте с закрытыми глазами в ЭЭГ человека, как правило, преобладают альфа-волны, хотя встречаются индивиды, у которых альфа-ритм почти не регистрируется. Общий рисунок, или паттерн, ЭЭГ каждого человека удивительно своеобразен. Его индивидуально-специфический характер, сложившись к 15-18 годам, сохраняется на протяжении длительного периода жизни, почти не меняясь. Эта особенность ЭЭГ побудила исследователей уже на первых этапах развития электроэнцефалографии искать наследственные причины наблюдающихся индивидуальных различий. Первое такое исследование было выполнено на 8 парах МЗ близнецов в 1936 г. (Davis H., Davis P., 1936). Авторы отметили поразительное сходство рисунков ЭЭГ у идентичных близнецов. Казалось, что у МЗ близнецов энцефалограммы отличаются не более чем у одного человека. Даже у МЗ близнецов, разлученных с самого раннего детства, паттерны ЭЭГ были удивительно похожими (Juel-Nielsen N., Harvald B., 1958). ЭЭГ-паттерн, как одна из надежных характеристик фенотипа, может быть использован для определения зиготности близнецов. Примеры энцефалограмм МЗ и ДЗ близнецов приведены на рисунке 10.3 а, б.
Генетические исследования ЭЭГ можно проводить, используя два разных подхода. Первый из них предполагает работу с целостным паттерном ЭЭГ. В этом случае ЭЭГ выступает как качественный признак. Сходство родственников при этом оценивается по принципу конкордантности. Исследователи пользуются при этом так называемым методом "слепой классификации". Это напоминает игру в парные картинки. После регистрации ЭЭГ у группы родственников (например, близнецов) приглашается независимый эксперт, который получает в свое распоряжение множество анонимных записей биоэлектрической активности. Его задача - подобрать пары по принципу сходства общего рисунка ЭЭГ. Те "картинки", которые он сочтет сходными, будут означать конкордантные пары, остальные - дискордантные. Таким способом выполнялись первые исследования. Наиболее последовательно это направление представлено в работах Ф. Фогеля и его коллег (Vogel F., 1970; Vogel F. et al., 1979). На этих исследованиях стоит остановиться подробнее. Для выяснения роли наследственности в вариативности ЭЭГ авторы пользовались близнецовым, семейным и популяционным методами. На больших контингентах испытуемых было выявлено 6 своеобразных паттернов ЭЭГ (рис. 10.4), которые довольно редко встречаются в популяции (до 5 %). При исследовании более 200 семей пробандов, т.е. лиц, обладающих этими вариантами ЭЭГ, оказалось, что почти все эти ЭЭГ-паттерны наследуются по аутосомно-доминантному типу (табл. 10.2). Три из них были впоследствии внесены в каталог "Наследственные признаки человека".

Таблица 10.2
Наследуемые ЭЭГ-варианты по Фогелю
Варианты ЭЭГ |
Количество сибсов |
Частота встречаемости в популяции (%) |
Типы наследования |
Низковольтная Низковольтная (пограничный вариант) |
117 |
4,2-4,6 2,1-2,3 |
Аутосомный-доминантный В немногих случаях аутосомный-доминантный, для остальных, видимо, крайний вариант обычной a-ЭЭГ |
Затылочные медленные B-волны (быстрый a-вариант, 16-19 кол/с) |
94 |
0,4-0,6 |
В основном аутосомный-доминантный |
Монотонные a-волны |
87 |
3,8-4,3 |
В основном аутосомный-доминантный. Некоторые трудности в классификации у молодых женщин |
Фронто-прецентальные B-группы |
65 |
0,4-1,5 |
Аутосомный-доминантный. Некоторые трудности в классификации у женщин |
Диффузорные B-волны |
103 |
3,3-4,0 |
Полигенный (с пороговым эффектом); у женщин более обычен, чем у мужчин; частота встречаемости увеличивается с возрастом |
Еще одним интересным моментом в исследованиях Ф. Фогеля является сопоставление психологических типов пробандов, обладающих описанными вариантами ЭЭГ. Психологическое обследование пробандов включало стандартные тесты, измеряющие интеллект, концентрацию внимания, особенности сенсорной и моторной сферы, а также стандартные личностные опросники. Приведем два описания для крайних вариантов ЭЭГ ("монотонные альфа-волны" и "низковольтная ЭЭГ"): 1. Монотонные альфа-волны (45 пробандов, аутосомно-доминантный тип наследования). Обладатель этого варианта ЭЭГ активен, стеничен, эмоционально стабилен, хорошо контролируем, устойчив к стрессу. Точность работы в тестах на концентрацию внимания и кратковременную память выше среднего, в то же время он не отличается высокой скоростью работы. Исследования семей выявили некоторую положительную ассортативность для этого типа ЭЭГ. Предполагаемая нейрофизиологическая основа указанных психологических особенностей - возможность значительной модуляции и селективного усиления афферентных стимулов благодаря особой регулярности альфа-ритма. (Предполагается, что одной из функций альфа-ритма ЭЭГ в приеме и переработке поступающих сигналов является скрининг и селективное усиление информации). 2. Низковольтная ЭЭГ (контртип предыдущего, 47 пробандов, аутосомно-доминантный тип наследования). Психологически пробанд отличается расслабленной и беззаботной позицией, экстраверт, ориентирован на группу, малоинициативен. Интеллект и особенно пространственные способности выше среднего. Относительно низкие показатели в тестах на концентрацию. Нейрофизиологически такой паттерн ЭЭГ должен быть связан со слабой модуляцией и селективным усилением афферентных стимулов. В дальнейших исследованиях пробандов, обладающих этими крайними вариантами ЭЭГ (Vogel F., Propping, 1981), были получены доказательства существования и биохимических различий. Изучалась активность фермента допамин-бета-гидроксилазы (ДБГ), причастного к синтезу одного из переносчиков нервного возбуждения в нервной системе. Средняя активность ДБГ у обладателей монотонных альфа-волн была почти вдвое выше, чем у обладателей противоположного типа (низковольтной ЭЭГ). Таким образом, наследственные различия в ЭЭГ оказались связанными и с различиями более высокого уровня (психологическими), и с различиями в биохимических процессах, которые ближе всего отстоят от уровня действия генов. Исследования такого рода демонстрируют важность изучения наследуемости не только психологических характеристик, но и сопоставления их с психофизиологическими и биохимическими показателями, являющимися промежуточными характеристиками фенотипа на пути от гена к поведению. Рассматривать ЭЭГ как качественный признак, естественно, можно, но вряд ли это достаточно перспективно, поскольку между выделенными Ф. Фогелем крайними вариантами паттернов существует множество переходных, не поддающихся четкой классификации. Случай с наследованием паттернов ЭЭГ является, по-видимому, довольно типичным для генетики поведения явлением. Сложный мультифакториальный признак варьирует столь значительно, что его можно рассматривать как результат проявления различных сочетаний множества генов. Возможно, существуют главные гены, определяющие какие-то крайние варианты признака, и тогда индивиды, у которых модифицирующие влияния других генов выражены слабее, будут демонстрировать наследование по Менделю, тогда как в остальных случаях мы будем иметь более сложное наследование. Ряд наследственных мультифакториальных заболеваний демонстрируют похожую картину. Например, для болезни Альцгеймера с ранним началом заболевания уже удалось обнаружить главные гены-кандидаты, тогда как все другие варианты заболевания пока еще исследуются. Исследования Ф. Фогеля представляют один из подходов к изучению генетики ЭЭГ. Здесь ЭЭГ рассматривается как качественный признак. Второй, более распространенный, подход связан с представлением ЭЭГ в виде ряда количественных показателей. В зависимости от своих возможностей исследователи измеряют различные параметры - частоты и амплитуды отдельных ритмических составляющих, спектральные характеристики, латентные периоды реакций, параметры синхронности ритмов в отдельных областях регистрации и т.д. Познакомиться с методами количественного анализа ЭЭГ можно в специальных руководствах по электроэнцефалографии. Набор таких количественных показателей характеризует каждую конкретную энцефалограмму. Соответственно, по каждому из параметров можно сравнивать родственников (близнецов, сибсов) и получать корреляции и оценки наследуемости. Задача изучения наследственности ЭЭГ напоминает задачу изучения наследственности почерка человека. Можно сопоставлять почерки на глаз, делая субъективные заключения о конкордантности и дискордантности родственников, а можно подойти более научно: разработать систему количественных оценок и проводить, например, измерения размеров букв, их наклона, количества букв в строке, расстояния между строками и т.д. В этом случае почерк каждого человека будет представлен в виде ряда количественных характеристик, по которым можно проводить сравнение родственников. Большинство исследователей, изучающих наследственность ЭЭГ, используют именно такой количественный подход. Кстати, в работах Ф. Фогеля такой способ оценки также представлен. На рисунке 10.5 приводится пример такого подхода с использованием метода близнецов (Мешкова Т.А., 1988). Столбцы диаграмм соответствуют коэффициентам внутриклассовой корреляции МЗ и ДЗ близнецов и пар неродственников по количественным параметрам альфа-ритма. Обращают на себя внимание очень высокие коэффициенты корреляции, характерные для МЗ близнецов. Коэффициенты корреляции ДЗ примерно наполовину ниже. Такое соотношение типично для аддитивного типа наследования количественного признака. Действительно, проверка гипотезы с помощью метода подбора моделей (см. тему 5) подтверждает высокий вклад аддитивной генетической составляющей (табл. 10.3). Вклад общей среды (VC) имеет место только в отведениях ЭЭГ с височных, теменных и центральных отделов мозга, во всех отведениях наблюдается больший или меньший вклад различающейся среды (VW).
Таблица 10.3
Коэффициенты внутриклассовой корреляции и разложение (в %) фенотипической дисперсии амплитуды альфа-ритма
Отведения |
rМЗ |
rДЗ |
VA |
VD |
VW |
VC |
x2 |
p |
T4 |
0,85 |
0,44 |
78 |
- |
15 |
7 |
6,8 |
<0,01 |
T3 |
0,64 |
0,50 |
57 |
- |
19 |
24 |
1,11 |
0,25-0,50 |
F4 |
0,95 |
0,45 |
95 |
- |
5 |
- |
4,06 |
0,10-0,20 |
F3 |
0,96 |
0,39 |
96 |
- |
4 |
- |
3,1 |
0,10-0,25 |
P4 |
0,92 |
0,61 |
60 |
- |
7,5 |
32,5 |
0,62 |
0,25-0,50 |
P3 |
0,95 |
0,51 |
85 |
- |
4,5 |
10,5 |
0,91 |
0,25-0,50 |
O2 |
0,96 |
0,33 |
95 |
- |
5 |
- |
1,94 |
0,25-0,50 |
O1 |
0,97 |
0,34 |
96 |
- |
4 |
- |
3,40 |
0,1-0,25 |
C4</SUB< td> |
0,88 |
0,49 |
60 |
- |
19 |
21 |
0,98 |
0,25-0,50 |
C3 |
0,82 |
0,40 |
82 |
- |
16,5 |
1,5 |
0,56 |
0,25-0,50 |
Недавно исследователи из Нидерландов (Beijsterveldt van C.E.M., Baal van G.C.M., 2002) подвели итог результатам работ по генетике ЭЭГ, выполненных в разные годы в различных странах. Во всех этих работах (всего их более 40) использовался количественный подход к анализу ЭЭГ (в основном спектральный анализ). Подавляющее большинство исследований, вошедших в обзор, были выполнены методом близнецов, хотя представлен и семейный метод. Авторами была применена процедура мета-анализа. Мета-анализ - это метод статистического анализа, позволяющий объединять результаты многочисленных самостоятельных исследований. Исходя из результатов 11 близнецовых исследований, выполненных в разных странах, авторы подсчитали, что в среднем показатель наследуемости спектральной мощности альфа-ритма составляет 79%. Это весьма высокий показатель. Для частоты альфа-ритма по результатам 5 близнецовых исследований "мета"-наследуемость частоты альфа-ритма оказалась также чрезвычайно высокой - 81% . На рисунках 10.6 и 10.7 приводятся результаты мета-анализа по параметрам альфа-ритма. Исследования наследуемости электроэнцефалограммы представляют интерес как своеобразный промежуточный этап к познанию механизмов наследуемости психологических признаков и различных психических заболеваний. Благодаря высокой наследуемости ЭЭГ может рассматриваться как один из перспективных маркеров для проведения анализа сцепления и анализа ассоциаций при изучении поведенческих фенотипов более высокого уровня.
В настоящее время уже удалось локализовать ген, ответственный за низкоамплитудный вариант ЭЭГ. Он оказался связан с маркером дистальной части длинного плеча хромосомы 20. Исследования вызванных потенциалов в психогенетике. Вызванные потенциалы (ВП) представляют собой изменения биоэлектрической активности в ответ на внешние или внутренние стимулы (как правило, в интервале 0-500 мсек после действия стимула). Эти колебания биоэлектрической активности называют также потенциалами мозга, связанными с событиями, поскольку они могут регистрироваться и при отсутствии раздражителя: если в цепи событий вероятность появления сигнала высока, но очередной сигнал не появляется, возникает биоэлектрический ответ на его отсутствие. ВП могут быть зарегистрированы и перед произвольным движением. Вызванные потенциалы несколько варьируют от стимула к стимулу, поэтому для их анализа сначала производят суммирование и усреднение отрезков ЭЭГ, сопутствующих событию. В дальнейшем анализируют усредненные колебания (рис. 10.8). ВП представляют собой ряд позитивных и негативных колебаний биопотенциала, которые возникают через короткий промежуток времени после подачи стимула, называемый латентным периодом (рис. 10.9). Вызванные потенциалы, как и ЭЭГ, подвергаются количественному анализу. Как правило, измеряются латентные периоды и амплитуды отдельных компонентов. Для удобства позитивные компоненты обозначают буквами Р, а негативные - N. Компоненты нумеруют в порядке появления (например, N1, P2) или в соответствии с латентным периодом его появления. Например, компонент Р300 - это позитивное колебание с латентным периодом 300 мсек.

Вызванные потенциалы отражают процессы переработки информации в нервной системе. Условно выделяют "экзогенные" и "эндогенные" компоненты. Экзогенные компоненты регистрируются сразу же после подачи стимула и отражают активность специфических проводящих путей, по которым поступают афферентные сигналы, и зон мозга, в которых они обрабатываются. Эндогенные компоненты связывают с активностью неспецифических ассоциативных областей коры. ВП, связанные с разными сенсорными системами (зрительные, слуховые, соматосенсорные, связанные с движениями) отличаются своими характеристиками (рис. 10.10). Для психофизиологии наибольший интерес представляют те колебания биоэлектрической активности, которые регистрируются позже 60 мсек, поскольку они отражают активность коры головного мозга и связаны с психологическими особенностями человека.
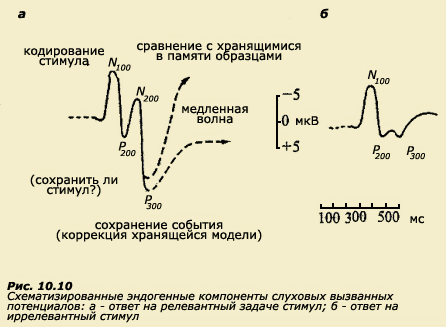
Вызванные
потенциалы, так же как ЭЭГ, отличаются
индивидуальным своеобразием. Индивидуальный
рисунок ВП также характеризуется
устойчивостью, т.е. мало варьирует при
повторных регистрациях у одного и того
же человека, хотя его устойчивость
меньше, чем ЭЭГ. Чтобы проверить, не
отражает ли индивидуальная специфика
ВП влияния наследственных факторов,
были проведены исследования близнецов
и других родственников. По многочисленности
эти исследования сильно уступают
психогенетическим исследованиям
интеллекта и других психологических
характеристик, но все же их достаточно,
чтобы можно было на сегодняшний день
сделать некоторые обобщающие
заключения.
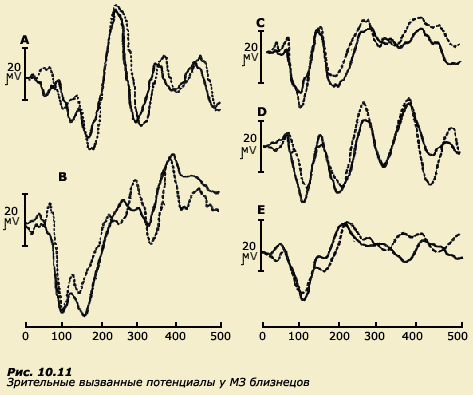
 Регистрация
вызванных потенциалов у близнецов
демонстрирует сходство волновой формы
у МЗ и меньшее совпадение у ДЗ (рис.
10.11, 10.12,
10.13, 10.14
и 10.17).
Регистрация
вызванных потенциалов у близнецов
демонстрирует сходство волновой формы
у МЗ и меньшее совпадение у ДЗ (рис.
10.11, 10.12,
10.13, 10.14
и 10.17).
В каждой паре ВП одного близнеца представлен сплошной линией, второго - пунктирной. Отметьте сходство латентностей пиков, волновой формы и изменений по мере роста интенсивности стимула. В паре 1 - слева - компонент P100N140 заметно возрастает по мере роста интенсивности стимула (augmenting) в паре 2 тот же компонент уменьшается (reducing).
Как правило, в исследованиях ВП применяют количественный подход и сходство родственников оценивают по параметрам латентных периодов и амплитуд отдельных компонентов, хотя в первых исследованиях оценивалось в основном общее сходство волновой формы. В уже упоминавшемся обзорном исследовании, выполненном недавно в Нидерландах (Beijsterveldt van C.E.M., Baal van G.C.M., 2002), помимо данных по ЭЭГ, авторы обобщили результаты генетических исследований вызванных потенциалов (около 30 работ). Среди всех характеристик ВП авторы выбрали для мета-анализа амплитуды и латентности эндогенного компонента Р300, поскольку в большинстве близнецовых исследований показатели наследуемости именно этого компонента варьируют от умеренных до высоких значений. Мета-наследуемость Р300 (по результатам пяти близнецовых исследований) оказалась равной в среднем 60% для амплитуды и 51% - для латентности (рис. 10.15).
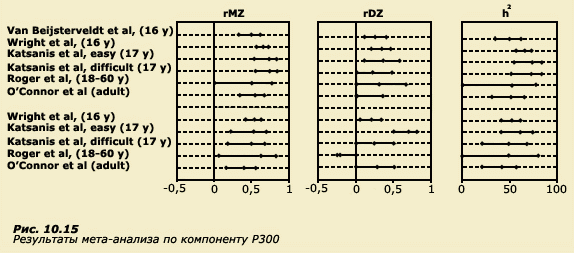
Как ЭЭГ, так и ВП позволяют изучать различия в биоэлектрической активности отдельных областей коры головного мозга, поскольку отведение биопотенциалов может осуществляться с различных точек поверхности головы. Это еще одно из преимуществ таких "эндофенотипов", как ЭЭГ и ВП. При изучении наследуемости биоэлектрических параметров часто отмечаются межзональные различия в коэффициентах кореляций между родственниками (в основном это близнецы) и в коэффициентах наследуемости. Приведем два примера. В исследовании ЭЭГ у близнецов, проведенном Т.А. Мешковой (1988), были обнаружены межполушарные различия в коэффициентах наследуемости, особенно характерные для височных зон коры. Коэффициенты наследуемости некоторых количественных параметров ЭЭГ оказались заметно более низкими в левом височном отведении по сравнению с правым. Возможно, это может объясняться более высокой пластичностью левого полушария, в котором у большинства людей локализуются речевые функции и моторика ведущей руки. Средовые воздействия, испытываемые в онтогенезе, могут оказывать большее влияние на структуры левого полушария, поскольку в процессе развития ребенка происходит интенсивное становление речи и праворукости. Топографические различия в коэффициентах наследуемости ВП были продемонстрированы в работе О'Коннера с соавторами (O'Conner S. et al., 1994; цит. по: Равич-Щербо И.В. с соавт., 1999). Схема эксперимента, которой в настоящее время широко пользуются нейрофизиологи, носит название oddball paradigm. От испытуемого требуется нажимать на кнопку при появлении редко встречающегося звука и игнорировать часто встречающийся. Такая схема позволяет исследовать избирательное (селективное) внимание. По современным представлениям процессы переработки информации и принятия решения при селективном внимании отражаются в компонентах N1 и Р300. Их амплитуды и латентности отличаются индивидуальной специфичностью и стабильностью. Выполненное на близнецах исследование обнаружило топографические различия в коэффициентах наследуемости (рис. 10.16).
Психогенетические исследования, проводимые с помощью метода вызванных потенциалов, позволяют взглянуть на особенности переработки информации в нервной системе в несколько ином ракурсе, чем это обычно делается в работах нейрофизиологов, когда не стоит задача оценки наследуемости. В качестве примера рассмотрим исследование зрительных ВП, проведенное на взрослых близнецах Т.М. Марютиной и Т.Г. Ивошиной (1984). Исследование интересно тем, что в нем использовалось семь вариантов зрительных стимулов: вспышка, симметричная геометрическая фигура, комбинация букв ДМО, хаотический набор элементов, из которых складывалось изображение дома, слово ДОМ, рисунок дома, шахматное поле. Следует обратить внимание на то, что ряд стимулов несли в себе семантическую нагрузку (рис. 10.17).
Авторам удалось получить интересные результаты: максимум наследственных влияний обнаружился в реакциях на вспышку и шахматное поле (60 и 62% соответственно), минимум - в ответах на семантические стимулы (рисунок дома и слово ДОМ - 31 и 29%). При разложении фенотипической дисперсии на наследственные и средовые компоненты, оказалось, что в реакциях на семантические стимулы значительный вклад в изменчивость вносит общая среда. Это исследование демонстрирует зависимость коэффициента наследуемости от условий эксперимента: наследственные различия ярче всего проявляются в физиологических реакциях на элементарные стимулы, не несущие смыслового содержания. При переработке смысловой информации на первый план выдвигаются средовые компоненты, за счет действия которых, по-видимому, формируется основная часть изменчивости зрительных вызванных потенциалов. Еще одно интересное психогенетическое исследование также связано с использованием метода вызванных потенциалов. С.Б. Малых (1986) регистрировал у близнецов потенциалы мозга, связанные с движением (ПМСД), в трех экспериментальных ситуациях. Первая ситуация требовала от испытуемого произвольно время от времени нажимать на кнопку. Во второй и третьей ситуациях то же самое движение испытуемого использовалось для сообщения о прогнозируемом событии. Сигналами служили звуки, подаваемые в наушники справа и слева, с разной вероятностью (1,0 и 0,5). Испытуемый нажатием на кнопку должен был предсказывать появление очередного звука в правом или левом наушнике. Регистрировались ПМСД, сопутствующие нажатию на кнопку. Оказалось, что вклад наследственных и средовых компонентов в вариативность показателей ПМСД, зарегистрированных в ответ на однотипное движение (нажатие на кнопку) существенно различается в зависимости от ситуации эксперимента. Чаще всего влияние генотипа обнаруживалось в ситуации прогноза равновероятностных событий (54% всех анализируемых показателей) и реже всего - при простых произвольных движениях (20% показателей). Поразительно, что при регистрации биоэлектрических реакций мозга, сопутствующих одному и тому же движению, мы наблюдаем разные причины межиндивидуальной вариативности. В первом случае гораздо больше вклад генетической составляющей, а во втором случае - средовой. По-видимому, как считает автор, все дело в том, что одно и то же движение оказалось в разном психологическом контексте. При произвольном нажатии на кнопку движение являлось целью деятельности испытуемого, а в ситуации прогноза целью был сам прогноз, а то же самое движение служило лишь средством для осуществления этой цели.
10.3. Двигательные характеристики
Так же как и в случае с восприятием или электроэнцефалограммой, в сфере движений есть некоторые варианты фенотипов, которые демонстрируют альтернативный характер изменчивости. Есть движения, которыми одни люди владеют в совершенстве, а другие - сколько бы не старались, овладеть не могут. Например, некоторые из представителей Homo sapiens могут шевелить ушами или сворачивать язык трубочкой, а у других такая способность отсутствует. Связано ли это с наследственными причинами и если да, то можно ли определить характер наследуемости? Может быть, это признаки, которые, подобно вкусовой чувствительности к некоторым веществам, наследуются по Менделю? Некоторые сведения на этот счет можно найти в интернет-каталоге наследственных признаков. Так, способность двигать ушами обнаруживается почти у 20% мужчин и 9,5% женщин. В 1949 г. Л. Линдер, сравнивая тех индивидов, которые умеют двигать ушами, с их родителями и сибсами, предположил, что эта способность, по-видимому, является наследственным признаком, проявляющим тенденцию к доминированию. Других специальных исследований мы пока не знаем. Внимание исследователей привлекли также некоторые способности, связанные с умением манипулировать языком: умение завивать кончик языка, складывать язык в форме трилистника (cloverleaf tongue, trefoil tongue), сворачивать язык трубочкой. Имеются данные, что способностью сворачивать язык обладают более 60% людей. В 1940 г. А. Стертевант описал два типа людей - "сворачивающие" и "несворачивающие" и сообщил, что способность к сворачиванию языка является доминантным признаком. Однако позже обнаружилось, что у МЗ близнецов наблюдается высокий процент дискордантности по этому признаку. По-видимому, способность сворачивать язык не связана с другой способностью - умением завивать кончик языка, однако может быть связана с умением двигать ушами. Редкая способность складывать язык в форме трилистника была описана в некоторых семьях и возможно наследуется доминантно. В целом нужно отметить, что в вопросе о наследуемости способности манипулировать языком до сих пор нет полной ясности: работ очень мало, а их результаты противоречивы. Еще один признак, который может быть, на первый взгляд, отнесен к разряду альтернативных - это право- леворукость, или просто "рукость" (этот термин стал употребляться в последнее время в отечественной литературе как прямой перевод английского handedness). Эта особенность двигательной сферы человека изучена в психогенетике гораздо полнее. Сначала рассмотрим сам признак. Частота леворукости в среднем составляет 5%, однако колебания этого показателя довольно значительны (от 1 до 30%) в зависимости от конкретной популяции, пола, рода занятий и возраста. Среди народностей Севера левши встречаются намного чаще, чем в других популяциях. Например, среди коренных жителей Таймыра их около 35%. У младенцев предпочтение руки не обнаруживается, хотя наблюдаются некоторые асимметрии позы, связанные с преимущественным поворотом головы вправо. Есть данные, свидетельствующие о том, что пропорция левшей в популяции с возрастом имеет тенденцию снижаться. В некоторых популяциях отмечается довольно значительное уменьшение числа леворуких среди людей старческого возраста, что побудило проверить, не различаются ли правши и левши по продолжительности жизни. Выяснилось, что среди людей, погибших от несчастного случая, левшей оказывается больше, чем правшей. В среднем левши и люди со смешанной рукостью (амбидекстры) умирают раньше, чем правши. Предполагается, что левши и амбидекстры являются своеобразной группой риска в связи с возможностью скрытой нейропатологии. Вместе с тем на юге Швеции было обнаружено, что среди женщин, больных раком груди, левши встречаются достоверно реже, чем среди здоровых женщин. Одним из возможных объяснений может быть общее влияние гормональных факторов на становление ручной асимметрии и риск рака молочной железы. Есть указания на связь рукости с аутоиммунными заболеваниями. Как видим, асимметрия рук связана с определенными биологическими факторами. В то же время не исключено влияние социальных причин, поскольку в становлении моторики рук немаловажную роль играют процессы научения. Вопрос о происхождении право- леворукости постоянно вызывает интерес исследователей. Очевидно, в основе наблюдаемых различий могут лежать как наследственные, так и средовые причины, в том числе патология развития. В 1940 г. Д. Райф (Rife D.), исследуя близнецов, предположил, что рукость является не альтернативным, а мультифакториальным признаком. Дальнейшие, довольно многочисленные исследования наследуемости рукости давали неоднозначные результаты, поэтому до сих пор нет единого взгляда на наследование правшества-левшества. Значительное место среди всех исследований в этой области занимают работы английского психолога М. Аннетт. М. Аннетт выдвинула гипотезу о наследовании рукости, согласно которой праворукость наследуется как признак с неполной доминантностью (Annett M., 1995). Ген, отвечающий за праворукость, М. Аннетт назвала геном правого сдвига (rs+). Согласно гипотезе М. Аннетт, доминантные гомозиготы (rs+ rs+) являются правшами и имеют доминантное левое полушарие, что предполагает локализацию речевых функций в левом полушарии. Соответственно рецессивные гомозиготы (rs- rs-) будут левшами с локализацией речи в правом полушарии. Гетерозиготы (rs+ rs-) могут быть как правшами, так и левшами и иметь любую локализацию речи. Несмотря на то, что М. Аннетт предлагает простую диаллельную модель наследования рукости, сам признак она рассматривает как количественный. Ею был разработан и стандартизован специальный двигательный тест (Peg Moving Test), позволяющий оценивать количественно моторику правой и левой руки. Тест требует от испытуемого переставлять колышки из одних ячеек в другие поочередно правой и левой рукой. Разница в затраченном времени указывает на степень ручной асимметрии. Выраженные правши будут более умело действовать правой рукой и быстрее справятся с заданием, выполняя тест правой рукой. Выраженные левши - наоборот, тогда как амбидекстры будут выполнять тест примерно одинаково обеими руками. В целом, если протестировать большую группу, можно получить непрерывное распределение по степени ручной асимметрии. Помимо модели М. Аннетт, предлагались и другие варианты наследования рукости. Например, в 1972 г. Дж. Леви и Т. Нагилаки предложили 2-генную 4-аллельную модель, согласно которой асимметрия полушарий и асимметрия рук контролируются разными локусами. Различные варианты генетических моделей наследования рукости предлагались и другими авторами. Все они, в основном, предполагают однолокусное наследование (Хрестомат. 10.1). В последние годы (2002-2003) появились молекулярно-генетические исследования рукости, в которых делаются попытки найти конкретные локусы, отвечающие за асимметрию рук. Франкс с коллегами (2002, 2003), используя для оценки фенотипа Peg Moving Test M. Аннетт, провели анализ сцепления для локусов количественного признака (QTL) на выборке из 105 пар взрослых братьев. Авторы пришли к заключению, что хотя вариативность правшества-левшества возникает за счет комплекса различных причин, все же можно считать, что имеется, по крайней мере, один полиморфный локус, который оказывает генетическое влияние на этот признак и располагается, скорее всего, на хромосоме 2. В заключение можно привести любопытный исторический пример, заставляющий задуматься о природе правшества-левшества. На границе между Англией и Шотландией располагается необычный замок-крепость Ферниехирст, принадлежавший династии Керров. Замок удивителен тем, что в его архитектуре все продумано для удобства людей, пользующихся в основном левой рукой. Например, винтовые лестницы закручиваются против часовой стрелки, тогда как во всех остальных замках - по часовой, ограждения устроены так, чтобы при защите от нападения удобнее было пользоваться левой рукой и т.п. Эндрю Керр, основавший династию Ферниехирст в 1457 г., несомненно, был левшой. Это давало ему явные преимущества в битвах. Предполагают, что он приучал своих сыновей и слуг также использовать в сражениях левую руку, а те, в свою очередь, учили этому своих сыновей. Ассоциация между фамилией Керр и леворукостью настолько велика, что среди шотландцев имя Керр стало нарицательным для обозначения левши (Kerr-handed = левша). Бодмер и МакКи (1994) приводят ссылку на обследование, проведенное врачами, которые всех пациентов с фамилией Керр опрашивали с целью выявления левшества. Оказалось, что среди людей с фамилией Керр 29,5% левшей и амбидекстров, тогда как среди других семейств их только 11%.
Психогенетические исследования других особенностей двигательной сферы человека, которые, несомненно, следует относить к количественным мультифакториальным признакам, также не слишком многочисленны, весьма разнообразны по применяемым подходам и часто противоречивы. С.Б. Малых, которы в 1988 г. сделал попытку свести воедино имеющиеся на тот момент результаты, выделяет четыре группы работ в соответствии с изучаемыми фенотипами:
1) стандартизованные двигательные пробы, 2) сложные поведенческие навыки, 3) физиологические системы обеспечения мышечной деятельности, 4) нейрофизиологический уровень обеспечения движений.
Стандартизованные двигательные пробы представляют собой различные тесты, специально разработанные для измерения моторных навыков. Один из таких тестов уже упоминался, когда речь шла о наследовании рукости у человека. Обычно такие тесты измеряют скорость, выносливость, силу, координацию, ловкость. Существуют две группы тестов. Первые применяются в основном для измерения спортивных навыков, вторые чаще используются в психодиагностике. Среди последних наиболее распространенными являются теппинг-тест (измерение скорости постукивания) и измерение времени сенсомоторной реакции (время между подачей сенсорного сигнала и двигательной реакцией испытуемого). Результаты немногочисленных работ первой группы (спортивные тесты), выполненных в основном на близнецах, обобщены на рисунке 10.18.
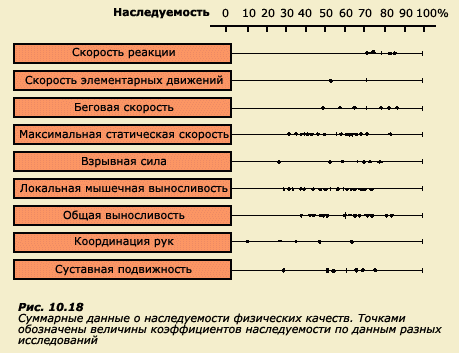
Можно видеть, что наиболее высокая наследуемость характерна для скоростных параметров, тогда как в тестах на координацию рук результаты сильно различаются, и большинство показателей наследуемости ниже 50%. Коэффициенты наследуемости, полученные в исследованиях времени реакции и теппинга, также весьма вариативны (от нулевых значений до чрезвычайно высоких - 70-90%). Учитывая немногочисленность исследований, чрезвычайное разнообразие методических схем экспериментов и возрастов испытуемых, следует заключить, что на характеристики двигательной деятельности, измеряемой с помощью специальных проб, действуют многочисленные факторы, которые не позволяют получить повторяющихся результатов. Единственное, что прослеживается в ряде работ, это больший вклад наследственности в вариативность двигательных характеристик, регистрируемых в условиях максимально возможной скорости работы (предельный темп деятельности). По-видимому, при высоких требованиях к скорости наследственные различия в двигательной деятельности проявляются более заметно. К сложным поведенческим навыкам относятся такие особенности, как походка и ходьба в целом, почерк, спортивные навыки, мимика и т.п. МЗ и ДЗ близнецы сравнивались по срокам начала ходьбы. Такие исследования проводились, в частности, в Москве в конце 20-х гг. в Медико-генетическом институте. Конкордантность МЗ близнецов по срокам начала хождения оказалась равной 67%, тогда как для ДЗ близнецов - лишь 29,9%. В других работах также имеются указания на более высокую конкордантность МЗ близнецов по срокам начала ходьбы. При проведении корреляционного анализа и вычислении наследственных и средовых составляющих фенотипической дисперсии оказалось, что довольно существенный вклад в сроки начала ходьбы вносит общесемейная среда (50-70%). В ряде работ у МЗ и ДЗ близнецов сравнивались особенности мимики и пантомимики. Как правило, исследователи указывают на высокое сходство мимических паттернов у МЗ близнецов при гораздо меньшем сходстве ДЗ близнецов (рис. 10.19). В отношении почерка близнецов, нет столь единодушного мнения. Еще Ф. Гальтон отметил, что почерки близнецов могут быть как похожими, так и непохожими (рис. 10.20).
Во всяком случае, почерки близнецов обычно не путают. Это же подтверждается и другими исследователями. Попытки количественно оценить детали почерка и сопоставить внутрипарное сходство близнецов по этим показателям не дали четких результатов. К сложным двигательным навыкам можно также отнести различные виды спорта. Известны семейные династии спортсменов. Среди известных спортсменов есть пары МЗ близнецов. Вообще МЗ близнецы характеризуются более высокой конкордантностью, чем ДЗ, по занятиям спортом. Вероятно, в склонности к спортивным занятиям наследственность играет не последнюю роль. Физиологическое обеспечение движений осуществляется системой органов дыхания, кровообращения, мышечным аппаратом, поэтому изучение вариативности показателей работы этих физиологических систем может быть также полезно при исследовании моторики в целом. Следует констатировать, что вопрос с точки зрения наследственности практически не изучен. Единственный показатель, по которому получены результаты на близнецах - это максимальное потребление кислорода (МПК) при выполнении спортивных движений. Относительно МПК следует отметить, что его среднепопуляционная величина составляет около 40 мл/мин/кг. Этнических различий по МПК не обнаруживается, показатель почти не изменяется с возрастом и мало подвержен тренировке, однако у спортсменов международного класса его величина почти в два раза выше среднепопуляционной. Исследования близнецов демонстрируют высокую наследуемость МПК (показатель наследуемости колеблется в пределах 66-93%). Вероятно, индекс МПК может быть использован как наследственный маркер для прогноза спортивной успешности. Психогенетические исследования потенциалов мозга, связанных с движениями, были описаны в предыдущем параграфе.
ВЫВОДЫ
10.1. Сенсорное восприятие
Одним из наиболее известных наследственных полиморфизмов человека, относящихся к сфере чувствительности, является ощущение вкуса фенилтиомочевины (ФТМ). Признак наследуется по Менделю. Обладатели доминантных генов (гомо- и гетерозиготы) ощущают горький вкус вещества, рецессивные гомозиготы считают ФТМ безвкусной.
Наследственные особенности обоняния, кинестетической и тактильной чувствительности практически не изучены.
Абсолютный музыкальный слух (способность безошибочно определять высоту звука) имеет тенденцию к накоплению в семьях, что позволяет предположить влияние наследственности на его формирование. Однако раннее начало занятий музыкой также способствует его развитию.
Нарушения цветового зрения (дальтонизм, дихромазии) контролируются сцепленными с полом рецессивными аллелями, поэтому чаще встречаются у мужчин, чем у женщин.
Некоторые особенности зрительного восприятия (иллюзии, критическая частота мельканий, время опознания изображения и т.п.) по данным различных авторов демонстрируют в основном коэффициенты наследуемости, превышающие 50%.
10.2. Морфология и физиология мозга
В строении и особенностях биоэлектрической активности мозга наблюдаются устойчивые межиндивидуальные различия. Отмечаемое своеобразие является результатом сложнейших взаимодействий наследственных и средовых факторов, а также многочисленных случайных событий, сопутствующих развитию нервной системы в онтогенезе.
Межиндивидуальная вариативность в объеме и размерах мозга человека определяется, очевидно, аддитивным генетическим компонентом (80-90%) и индивидуальной средой.
Некоторые редкие паттерны (своеобразные индивидуальные варианты рисунка) ЭЭГ наследуются по Менделю и коррелируют с психологическими характеристиками и биохимическими особенностями мозга.
Роль наследственности в вариативности альфа-ритма ЭЭГ человека по данным мета-анализа составляет порядка 80%. Благодаря высокой наследуемости ЭЭГ может рассматриваться как один из перспективных маркеров для проведения анализа сцепления и анализа ассоциаций при изучении фенотипов более высокого уровня.
Мета-наследуемость компонента Р300 вызванных потенциалов (ВП) человека (по результатам пяти близнецовых исследований) составляет 50-60%.
В коэффициентах наследуемости количественных характеристик ЭЭГ и ВП наблюдаются возрастные, межзональные, кроссситуативные и другие различия. Особого внимания заслуживают изменения наследуемости характеристик ВП, связанные со спецификой психологической структуры деятельности испытуемого.