

3.5. Гены в популяциях. Закон Харди-Вайнберга Формулировка закона Харди-Вайнберга заключается в следующем:
При определенных условиях (замкнутость популяции, отсутствие отбора и случайность слияния гамет) частоты аллей сохраняются из поколения в поколение.
Частоты встречаемости гомо- и гетерозигот устанавливаются за одно поколение и соответствуют отношению p2: 2pq : q2.
Словарь терминов
Хромосома
Сперматозоид
Моногибридное скрещивание
Дигибридное скрещивание
Гамета
Рецессивный
Доминантный
Рекомбинация
Кроссинговер
Гомозигота
Гетерозигота
Вопросы для самопроверки
Каковы были представления о наследственности до работ Г. Менделя?
В чем заключается революционный характер открытия Г. Менделя?
Как можно доказать, что наследственные факторы дискретны и парны?
Какие выводы были сделаны Г. Менделем на основании расщепления при моногибридном скрещивании?
Что такое доминантный и рецессивный признаки?
Что такое решетка Пеннета? Как выглядит решетка Пеннета для моногибридного скрещивания?
Что такое дигибридное скрещивание и какой вид при этом имеет расщепление?
Как выглядит решетка Пеннета для дигибридного скрещивания?
Как можно представить себе дигибридное скрещивание у человека?
Что такое рекомбинация и при образовании каких клеток она происходит?
Что такое хромосома?
На основании чего было сделано предположение о связи дискретных единиц наследственности с хромосомами?
Что такое диплоидный и гаплоидный наборы хромосом?
В каких клетках человека имеется гаплоидный набор хромосом?
Раздел III. Биометрическая генетика глава 4. Генетические основы количественной изменчивости
4.1. Количественная изменчивость и методы ее описания
4.2. Наследственность и среда как факторы возникновения количественной изменчивости
4.3. Показатель наследуемости и его особенности
4.4. Генотип-средовое взаимодействие
Во второй теме мы кратко описали путь менделевской генетики с момента открытия дискретных единиц наследственности и до познания молекулярной природы гена. Мы специально ограничивались рассмотрением качественных признаков, поскольку механизмы их наследуемости и изменчивости могут быть представлены в виде достаточно простых схем. Признаки, для которых характерна дискретная изменчивость, обычно контролируются одним-двумя главными генами, у которых может быть два или несколько аллелей, а внешние условия относительно мало или почти совсем не влияют на степень их проявления. Напомним еще раз примеры таких признаков у человека. Это различные группы крови, цветовая слепота, некоторые наследственные аномалии. Не так просто обстоит дело с количественными, континуальными признаками, обладающими непрерывной изменчивостью, такими, как рост, вес, цвет кожи и т.п. Напомним, что большинство интересующих нас психологических характеристик (интеллект, темперамент и т.д.) относится именно к этой категории признаков, и сейчас мы переходим к рассмотрению вопросов, связанных с генетикой количественных признаков.
4.1. Количественная изменчивость и методы ее описания
4.1.1. Измерение количественных признаков
4.1.2. Характеристики центральной тенденции
4.1.3. Характеристики разброса
4.1.4. Межгрупповые и межиндивидуальные различия
4.1.1. Измерение количественных признаков
Итак, мы уже знаем
о существовании двух типов изменчивости.
Дискретная, или качественная, изменчивость
ограничивается рядом четко выраженных
признаков, не имеющих промежуточных
форм. Непрерывная, или количественная,
изменчивость, предполагает существование
непрерывного ряда переходов от минимальной
выраженности признака до максимальной
(рис. 2.1 и 2.2).
Прежде
чем перейти к анализу факторов, приводящих
к возникновению непрерывной изменчивости,
необходимо остановиться на методах,
которые применяются для описания этой
изменчивости, и рассмотреть основные
статистические понятия, с которыми
работает количественная
генетика, и генетика
поведения в
частности.
Любой
количественный признак можно измерять
с разной точностью, в зависимости от
поставленной задачи и того измерительного
инструмента, которым исследователь
располагает. Рост можно измерить с
точностью до сантиметра, вес - с точностью
до грамма, но в последнем случае, пожалуй,
ошибка измерения перекроет указанную
точность, и такое измерение веса окажется
просто бессмысленным. В психологии
также существуют измерительные
инструменты, чаще всего тесты или
опросники, и исследователь сам решает,
что и как измерять и с какой точностью.
Все требования к психологическим
измерениям в генетике поведения остаются
теми же, которым должны удовлетворять
психодиагностические процедуры -
надежность, валидность,
репрезентативность (подробнее об этом
можно узнать из руководств по
психодиагностике). После того как выбран
психологический конструкт, с которым
работает исследователь, выбраны методы
измерения и спланирована выборка
исследования, можно приступать к реальным
измерениям. Что мы получим в результате?
Конечно же, тот самый ряд непрерывной
изменчивости, о котором мы говорили
ранее. Этот ряд можно представить в виде
распределения частот встречаемости
различных величин изучавшегося
признака.
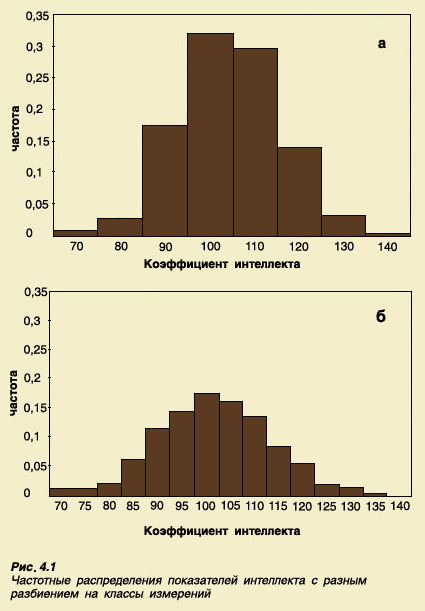 Например,
проводится измерение интеллекта (в
баллах IQ - коэффициента интеллекта) у
детей 10 лет в популяции жителей некоего
города. Скорее всего, окажется, что
небольшая часть детей будет иметь
относительно низкие баллы интеллекта
- 70-80 баллов; какая-то, тоже небольшая,
часть - очень высокие баллы - 120-130 и выше.
Основная же масса детей будет
характеризоваться средним интеллектом
в пределах от 90 до 110. Все показатели
интеллекта мы можем разбить на классы,
например, с шагом в 10 баллов, и представить
число детей, попавших в каждый класс
измерений, в виде диаграммы (рис. 4.1а).
Эта диаграмма представляет собой
распределение частот различных величин
IQ в обследованной группе. Такое
распределение частот отражает количество
детей, попавших в каждый класс измерений.
Если разбиение на классы сделать более
дробным - с шагом в 5 баллов, то распределение
будет таким, как показано на рисунке
4.1б. Поскольку в этом случае число классов
в два раза больше, чем в предыдущем,
число детей, попавших в каждый класс
измерений, будет меньше, чем в предыдущем
примере, и форма распределения изменится
- оно будет более плоским. Но если охватить
измерениями в два раза больше детей, мы
снова получим распределение, похожее
по форме на первое, но несколько более
сглаженное, напоминающее кривую
нормального распределения.
Теперь
предположим, что такие измерения
интеллекта мы провели в трех различных
группах детей. Одну группу пусть составят
дети, обучающиеся во вспомогательных
школах, вторую группу - дети, обучающиеся
в специальных школах для одаренных
детей, и третью - дети, обучающиеся в
массовых школах. Какого рода распределения
мы можем получить? Скорее всего, основная
масса детей из вспомогательных школ
будет иметь невысокие баллы интеллекта,
ниже среднего в популяции.
Одаренные дети будут, напротив,
характеризоваться более высоким
интеллектом; дети же из массовых школ,
вероятно, покажут результаты, близкие
к среднепопуляционным. На рисунке 4.2
представлены распределения, которые
соответствуют трем гипотетическим
группам обследованных. Как мы видим,
эти три частотных распределения
отличаются некоторыми особенностями.
Каковы же их количественные характеристики?
Например,
проводится измерение интеллекта (в
баллах IQ - коэффициента интеллекта) у
детей 10 лет в популяции жителей некоего
города. Скорее всего, окажется, что
небольшая часть детей будет иметь
относительно низкие баллы интеллекта
- 70-80 баллов; какая-то, тоже небольшая,
часть - очень высокие баллы - 120-130 и выше.
Основная же масса детей будет
характеризоваться средним интеллектом
в пределах от 90 до 110. Все показатели
интеллекта мы можем разбить на классы,
например, с шагом в 10 баллов, и представить
число детей, попавших в каждый класс
измерений, в виде диаграммы (рис. 4.1а).
Эта диаграмма представляет собой
распределение частот различных величин
IQ в обследованной группе. Такое
распределение частот отражает количество
детей, попавших в каждый класс измерений.
Если разбиение на классы сделать более
дробным - с шагом в 5 баллов, то распределение
будет таким, как показано на рисунке
4.1б. Поскольку в этом случае число классов
в два раза больше, чем в предыдущем,
число детей, попавших в каждый класс
измерений, будет меньше, чем в предыдущем
примере, и форма распределения изменится
- оно будет более плоским. Но если охватить
измерениями в два раза больше детей, мы
снова получим распределение, похожее
по форме на первое, но несколько более
сглаженное, напоминающее кривую
нормального распределения.
Теперь
предположим, что такие измерения
интеллекта мы провели в трех различных
группах детей. Одну группу пусть составят
дети, обучающиеся во вспомогательных
школах, вторую группу - дети, обучающиеся
в специальных школах для одаренных
детей, и третью - дети, обучающиеся в
массовых школах. Какого рода распределения
мы можем получить? Скорее всего, основная
масса детей из вспомогательных школ
будет иметь невысокие баллы интеллекта,
ниже среднего в популяции.
Одаренные дети будут, напротив,
характеризоваться более высоким
интеллектом; дети же из массовых школ,
вероятно, покажут результаты, близкие
к среднепопуляционным. На рисунке 4.2
представлены распределения, которые
соответствуют трем гипотетическим
группам обследованных. Как мы видим,
эти три частотных распределения
отличаются некоторыми особенностями.
Каковы же их количественные характеристики?
Во-первых, все три распределения по-разному расположены на шкале измерения, то есть они отличаются по своей центральной тенденции.
Во-вторых, они различаются по разбросу значений - если в двух отобранных группах (дети из вспомогательных школ и одаренные дети) разброс значений вокруг центральной тенденции невелик, то в группе неотобранных детей (массовые школы) разброс заметно выше.

4.1.2. Характеристики центральной тенденции
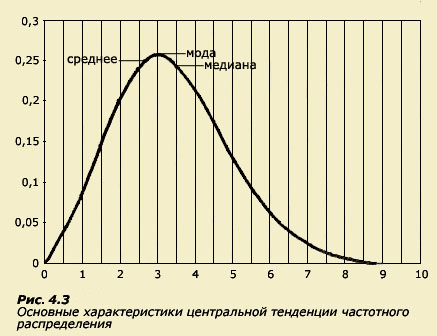
Существуют три меры центральной тенденции, характеризующие любое распределение. Их не следует смешивать, поскольку получаемые с их помощью оценки могут и не совпадать (рис. 4.3). Первая - это мода, или наиболее часто встречающееся значение признака. Мода соответствует вершине распределения. Вторая характеристика - медиана - представляет собой такое значение, выше и ниже которого располагаются результаты 50% людей. И, наконец, наиболее часто используемая и известная всем характеристика - это среднее, то есть среднее арифметическое, определяемое путем суммирования всех значений измерявшегося признака и деления полученной суммы на число обследованных. Для некоторых распределений мода, медиана и среднее различаются, для некоторых - совпадают (это так называемое нормальное распределение). Если распределение асимметрично, т.е. имеет длинный "хвост" с одной стороны, мода, медиана и среднее будут значительно отличаться.
4.1.3. Характеристики разброса
Для характеристики разброса значений вокруг среднего чаще всего пользуются показателем дисперсии. Дисперсия представляет собой среднее арифметическое квадратов разностей между наблюдаемыми значениями и средней величиной:
![]()
Если многие значения сильно отличаются от среднего, дисперсия будет высокой, а распределение растянутым. Если же значения признака у обследованных индивидов группируются вблизи средней величины, то дисперсия будет низкой. В нашем примере распределение оценок интеллекта у отстающих, и одаренных детей характеризуется примерно одинаковой невысокой дисперсией, и распределения отличаются лишь центральной тенденцией; третье распределение (дети из массовых школ) более растянуто и характеризуется более высокой дисперсией. Для описания разброса можно пользоваться и другой характеристикой - стандартным отклонением, величина которого равна корню квадратному из дисперсии.
4.1.4. Межгрупповые и межиндивидуальные различия
Можно проводить измерения в самых различных группах людей - у детей и взрослых, у мужчин и женщин, жителей городов и сельской местности, и всякий раз мы будем получать распределения, характеризующиеся средними и дисперсиями. Различия между получаемыми распределениями могут быть существенными или несущественными. Чаще всего в психологических исследованиях основное внимание уделяется проблеме межгрупповых различий, обусловленных полом, возрастом и т.п., и, как правило, основной величиной, с которой работают исследователи, является среднее значение изучаемой характеристики в каждой из групп. Генетику поведения больше интересуют не различия между группами, а различия между отдельными индивидами внутри группы, поэтому величина дисперсии, характеризующая величину различий в группе, представляет для психогенетика самостоятельную ценность. Весь математический аппарат современной генетики поведения рассчитан на работу с дисперсиями.
Выводы
Требования к психологическим измерениям в генетике поведения соответствуют основным требованиям психометрики (надежность, валидность, репрезентативность).
Распределение частот встречаемости различных количественных значений признака в популяции характеризуется двумя статистическими величинами - центральной тенденцией (мода, медиана, среднее) и разбросом значений вокруг среднего (дисперсия).
Дисперсия характеризует межиндивидуальные различия (изменчивость, вариативность).
Генетика поведения изучает природу индивидуальных различий.
4.2. Наследственность и среда как факторы возникновения количественной изменчивости
4.2.1. Генотип и фенотип
4.2.2. Возникновение количественной изменчивости под действием полимерных генов. Генетическая дисперсия
4.2.3. Различные типы взаимодействия генов Аддитивное взаимодействие
4.2.4. Возникновение количественной изменчивости под действием среды. Норма реакции. Средовая дисперсия
4.2.5. Совместный вклад генотипа и среды в количественную изменчивость. Генетическая и средовая дисперсии как составляющие популяционной фенотипической дисперсии
Как мы видели, существуют признаки, которые имеют простое однозначное соответствие генам (группы крови, например), и среда не оказывает никакого влияния на проявление таких признаков. Человек, обладающий аллелями группы крови А, при любых обстоятельствах будет иметь эту группу крови, в какой бы среде он ни находился. Но такие полностью генетически детерминированные признаки являются скорее исключением, чем правилом. Большинство же признаков человека есть результат взаимодействия генов с факторами среды. Признаки, для которых характерна непрерывная изменчивость, как правило, обусловлены совместным действием многих генов и факторов среды. Наследуемость таких признаков называют полигенной.
4.2.1. Генотип и фенотип
В генетике существуют два очень важных понятия. Это понятия генотип и фенотип. Мы уже знаем, что наследственная конституция складывается из большого числа различных генов. Вся совокупность генов данного организма называется его генотипом, то есть понятие генотипа идентично понятию генетической конституции. Свой генотип (набор генов) каждый человек получает в момент зачатия и несет его без всяких изменений через всю свою жизнь. Активность генов может меняться, но их состав остается неизменным. От понятия генотип следует отличать другое сходное понятие - геном. Геномом называется совокупность генов, характерная для гаплоидного набора хромосом особи данного вида. В отличие от генотипа геном является характеристикой вида, а не отдельной особи. Фенотип же представляет собой любые проявления организма в каждый момент его жизни. Фенотип включает в себя и внешний вид, и внутреннее строение, и физиологические реакции, и любые формы поведения, наблюдаемые в текущий момент. Например, уже упоминавшиеся группы крови системы АВ0 - это пример фенотипа на физиолого-биохимическом уровне. Хотя на первый взгляд многим кажется, что группа крови - это генотип, поскольку она четко определяется действием генов и не зависит от среды, однако это лишь проявление действия генов, и поэтому должно быть отнесено к категории фенотипов. Вспомним, что представители групп крови А или В могут иметь разные генотипы (гомозиготные и гетерозиготные). Сложными фенотипами являются все поведенческие проявления. Например, почерк, который отличает данного индивида, является его поведенческим проявлением и также относится к категории фенотипов. Если группа крови в течение жизни не меняется, то почерк по мере тренировки навыка письма претерпевает значительные изменения. Если генотипы наследуются и остаются неизменными в течение жизни индивида, то фенотипы большей частью не наследуются - они развиваются и являются следствием наших генотипов лишь в определенной мере, поскольку большую роль в становлении фенотипов играют условия внешней среды. Весь процесс развития от оплодотворенной яйцеклетки до взрослого организма происходит не только под непрерывным регулирующим влиянием генотипа, но и под влиянием множества различных условий среды, в которых находится растущий организм. Поэтому необычайная изменчивость, свойственная живым организмам, обусловлена не только огромным разнообразием генотипов, возникающим вследствие рекомбинации генов и мутационного процесса, но в значительной степени объясняется и тем, что отдельные индивиды развиваются в различающихся условиях среды. С давних пор идет полемика о том, что важнее для формирования организма - среда или генетическая конституция. Особенно острые споры разгораются там, где дело касается поведения человека, его психологических характеристик - темперамента, умственных способностей, черт личности. Не случайно, что именно с вопроса о природе умственной одаренности начались исследования в области генетики человека. Ф. Гальтон первым в научном трактате поставил рядом два понятия, которые в той или иной форме не сходят со страниц научной литературы до наших дней. Это понятия - "nature and nurture", то есть "природа и условия воспитания". Генетиков, и генетиков поведения в частности, часто упрекают в отрицании роли среды. Однако такой упрек совершенно необоснован. Одним из основных постулатов генетики является тезис о том, что фенотип представляет собой результат взаимодействия генотипа и среды. В процессе этого взаимодействия и возникает то многообразие фенотипических проявлений, которое характерно для большинства признаков человека, относящихся к категории количественных и образующих непрерывный ряд изменчивости.
4.2.2. Возникновение количественной изменчивости под действием полимерных генов. Генетическая дисперсия
Забудем на время
о влиянии факторов среды на фенотипические
проявления и остановимся только на
проблеме совместного действия многих
генов, которое само по себе приводит к
количественной изменчивости.
Мы
уже упоминали, что в первые годы развития
менделевской генетики основное внимание
было уделено изучению наследования
качественных признаков. Все попытки
проанализировать наследование
количественных признаков (высота стебля
у растений, размеры листьев, длина колоса
и т.п.) наталкивались на множество
затруднений, главным из которых была
невозможность различить разные категории
потомства. Изменчивость во втором
поколении (F2) мало чем отличалась от
изменчивости в первом поколении (F1).
Дело в том, что гибриды F1 по количественным
признакам занимают промежуточное
положение между родителями. Эта тенденция
сохраняется и в последующих
поколениях.
 Лишь
в 1910 г. было обнаружено, что количественные
признаки обусловлены точно такими же
расщепляющимися генами и их передача
также осуществляется в соответствии с
теми же менделевскими
законами, справедливыми
для качественных признаков. Это открытие,
связанное с именем Г. Нильссона-Эле,
создало прочный фундамент для дальнейшего
развития генетики количественных
признаков.
Г.
Нильссон-Эле изучал наследование окраски
зерен у пшеницы и овса и при дигибридном
расщеплении получал, в отличие от уже
знакомого нам соотношения 9 : 3 : 3 : 1,
особое соотношение - 15 : 1. 15/16 зерен были
окрашенными и лишь 1/16 - белыми. Такое
расщепление Г. Нильссон-Эле объяснил
сходным действием нескольких генов.
Такие гены называют полимерными.
Это означает, что два или более генов
вызывают развитие одного и того же
признака. Если мы вспомним решетку
Пеннета для дигибридного расщепления
(см.
табл. 3.3), то увидим, что во всех
клетках, кроме одной, обязательно
присутствует хотя бы один доминантный
аллель - только одна
комбинация является полностью рецессивной.
Отсюда становится ясным, почему
расщепление, полученное Г. Нильссоном-Эле,
соответствует отношению 15:1: пятнадцать
генотипов, содержащих доминантные
аллели, дали окрашенные зерна, и лишь
один генотип, представляющий собой
двойной рецессив,
дал белые зерна, по своему фенотипу
значительно отличающиеся от остальных.
Если полимерных генов не два, а три
(тригибридное расщепление), то решетка
будет иметь 64 квадрата, и опять только
один из них будет содержать полностью
рецессивный генотип. Соответственно
63 квадрата будут нести хотя бы один
доминантный аллель и расщепление
окажется 63:1. Такое расщепление также
получал Г. Нильссон-Эле в своих
экспериментах.
При
полимерном наследовании эффекты
доминантных генов могут быть двоякого
рода. В одних случаях оказывается
достаточным присутствия одного
доминантного аллеля, чтобы признак
максимально проявился - добавление
дополнительных доминантных аллелей не
оказывает заметного эффекта. В этом
случае возникают расщепления типа 15:1
или 63:1, в которых не бывает промежуточных
классов. Но существует и другой вариант
действия полимерных генов, когда эффект
тем больше, чем больше доминантных
аллелей в генотипе. Например, комбинация
Aabb будет давать более слабый эффект,
чем комбинация AAbb или комбинация
АаВb и т.д. Понятно, что наибольший
эффект будет у комбинации аллелей ААВВ,
а наименьший - у комбинации aabb.
Соответственно в этом случае при
гибридизации полностью доминантной
формы с формой, полностью рецессивной,
мы получим ряд промежуточных
классов.
В
случае действия множества генов
различные степени проявления признака
фактически обусловлены той же самой
дискретностью генетических факторов,
что и в опытах Менделя
с качественными признаками, однако
внешне эффект будет выражаться в
непрерывной изменчивости - чем больше
доминантных аллелей в генотипе, тем
степень выраженности признака сильнее,
и наоборот. Результатом совместного
действия генов на один и тот же признак
будет нормальное распределение признака
в популяции.
Более наглядно процесс возникновения
нормального распределения признака
можно продемонстрировать на
примере.
Возьмем
случай дигибридного скрещивания, то
есть пример действия двух независимых
генов, каждый из которых имеет пару
аллелей. Оба гена обусловливают развитие
одного и того же признака, эффекты
отдельных аллелей суммируются. Пусть
аллели A1 и А2 вызывают
усиление признака, а аллели a1 и
а2 - его ослабление. Теперь посмотрим,
какое потомство могут дать гетерозиготы
А1 а1 А2 а2. Заполним
решетку Пеннета для дигибридного
скрещивания (табл. 4.1).
Ц
Лишь
в 1910 г. было обнаружено, что количественные
признаки обусловлены точно такими же
расщепляющимися генами и их передача
также осуществляется в соответствии с
теми же менделевскими
законами, справедливыми
для качественных признаков. Это открытие,
связанное с именем Г. Нильссона-Эле,
создало прочный фундамент для дальнейшего
развития генетики количественных
признаков.
Г.
Нильссон-Эле изучал наследование окраски
зерен у пшеницы и овса и при дигибридном
расщеплении получал, в отличие от уже
знакомого нам соотношения 9 : 3 : 3 : 1,
особое соотношение - 15 : 1. 15/16 зерен были
окрашенными и лишь 1/16 - белыми. Такое
расщепление Г. Нильссон-Эле объяснил
сходным действием нескольких генов.
Такие гены называют полимерными.
Это означает, что два или более генов
вызывают развитие одного и того же
признака. Если мы вспомним решетку
Пеннета для дигибридного расщепления
(см.
табл. 3.3), то увидим, что во всех
клетках, кроме одной, обязательно
присутствует хотя бы один доминантный
аллель - только одна
комбинация является полностью рецессивной.
Отсюда становится ясным, почему
расщепление, полученное Г. Нильссоном-Эле,
соответствует отношению 15:1: пятнадцать
генотипов, содержащих доминантные
аллели, дали окрашенные зерна, и лишь
один генотип, представляющий собой
двойной рецессив,
дал белые зерна, по своему фенотипу
значительно отличающиеся от остальных.
Если полимерных генов не два, а три
(тригибридное расщепление), то решетка
будет иметь 64 квадрата, и опять только
один из них будет содержать полностью
рецессивный генотип. Соответственно
63 квадрата будут нести хотя бы один
доминантный аллель и расщепление
окажется 63:1. Такое расщепление также
получал Г. Нильссон-Эле в своих
экспериментах.
При
полимерном наследовании эффекты
доминантных генов могут быть двоякого
рода. В одних случаях оказывается
достаточным присутствия одного
доминантного аллеля, чтобы признак
максимально проявился - добавление
дополнительных доминантных аллелей не
оказывает заметного эффекта. В этом
случае возникают расщепления типа 15:1
или 63:1, в которых не бывает промежуточных
классов. Но существует и другой вариант
действия полимерных генов, когда эффект
тем больше, чем больше доминантных
аллелей в генотипе. Например, комбинация
Aabb будет давать более слабый эффект,
чем комбинация AAbb или комбинация
АаВb и т.д. Понятно, что наибольший
эффект будет у комбинации аллелей ААВВ,
а наименьший - у комбинации aabb.
Соответственно в этом случае при
гибридизации полностью доминантной
формы с формой, полностью рецессивной,
мы получим ряд промежуточных
классов.
В
случае действия множества генов
различные степени проявления признака
фактически обусловлены той же самой
дискретностью генетических факторов,
что и в опытах Менделя
с качественными признаками, однако
внешне эффект будет выражаться в
непрерывной изменчивости - чем больше
доминантных аллелей в генотипе, тем
степень выраженности признака сильнее,
и наоборот. Результатом совместного
действия генов на один и тот же признак
будет нормальное распределение признака
в популяции.
Более наглядно процесс возникновения
нормального распределения признака
можно продемонстрировать на
примере.
Возьмем
случай дигибридного скрещивания, то
есть пример действия двух независимых
генов, каждый из которых имеет пару
аллелей. Оба гена обусловливают развитие
одного и того же признака, эффекты
отдельных аллелей суммируются. Пусть
аллели A1 и А2 вызывают
усиление признака, а аллели a1 и
а2 - его ослабление. Теперь посмотрим,
какое потомство могут дать гетерозиготы
А1 а1 А2 а2. Заполним
решетку Пеннета для дигибридного
скрещивания (табл. 4.1).
Ц ифры,
проставленные в правом нижнем углу
каждой клетки, указывают на количество
аллелей,
усиливающих признак, в каждом генотипе.
Мы видим, что больше всего клеток
оказалось с цифрой 2. Представим это в
виде диаграммы, в которой в отдельных
столбцах объединены генотипы с одинаковым
количеством аллелей-усилителей
признака.
Полученная
нами диаграмма напоминает гистограмму
нормального распределения для пяти
градаций (классов) какого-либо признака.
На оси абсцисс обозначено количество
аллелей-усилителей, которому соответствует
определенная степень выраженности
признака, а на оси ординат - наблюдаемая
частота для каждого из классов. Не
вдаваясь в подробности, отметим, что
частоты 1:4:6:4:1 соответствуют коэффициентам
разложения бинома Ньютона (а+b)4.
При любом количестве пар генов n частоты
градаций количественного признака в
потомстве будут соответствовать
коэффициентам разложения бинома Ньютона
(а+b)n.
Мы
привели пример, в котором суммируются
эффекты всего двух двухаллельных генов.
Если таких генов будет три, то количество
классов увеличится до 7, а число возможных
комбинаций составит 64, для четырех генов
мы будем иметь 9 классов и 256 комбинаций
и т.д. Простой подсчет показывает, что
при увеличении числа локусов
и аллелей
с суммирующимся эффектом действия на
определенный признак число возможных
генотипов
нарастает очень быстро. Так, если
предположить, что гены какого-либо
количественного признака располагаются
в 20 локусах и имеют по 4 аллеля, то
количество генотипов будет составлять
1020. Понятно, что частотная
гистограмма степени выраженности
признака при таком количестве генотипов
будет все более сглаживаться и напоминать
нормальное распределение. Вся изменчивость
при этом будет носить чисто генетический
характер, и количественная величина,
характеризующая вариативность признака
в популяции (дисперсия), в данном случае
может быть названа генетической
дисперсией.
В рассмотренном примере вся фенотипическая
изменчивость объясняется наследственной
вариативностью, т.е. VP = VG. В
генетике для обозначения дисперсии
используется латинская буква V (от
английского variance). Значками VP VG
и VE обозначаются соответственно
фенотипическая (P = phenotype), генотипическая
(G = genotype) и средовая (Е = environment) дисперсии.
Однако
подобный же непрерывный характер может
носить и изменчивость, целиком определяемая
средовыми факторами. Поэтому, столкнувшись
с непрерывной изменчивостью, невозможно
сразу определить, обусловлена ли она
исключительно воздействиями среды или
здесь имеет место и полигенный
характер наследуемости. В генетике
существуют специальные методы, позволяющие
определить природу изменчивости, но об
этом речь пойдет немного позже. Сейчас
же только подчеркнем, что в большинстве
случаев непрерывный характер изменчивости
определяется как действием многих
генов, так и взаимодействием генотипа
со всевозможными средовыми условиями.
ифры,
проставленные в правом нижнем углу
каждой клетки, указывают на количество
аллелей,
усиливающих признак, в каждом генотипе.
Мы видим, что больше всего клеток
оказалось с цифрой 2. Представим это в
виде диаграммы, в которой в отдельных
столбцах объединены генотипы с одинаковым
количеством аллелей-усилителей
признака.
Полученная
нами диаграмма напоминает гистограмму
нормального распределения для пяти
градаций (классов) какого-либо признака.
На оси абсцисс обозначено количество
аллелей-усилителей, которому соответствует
определенная степень выраженности
признака, а на оси ординат - наблюдаемая
частота для каждого из классов. Не
вдаваясь в подробности, отметим, что
частоты 1:4:6:4:1 соответствуют коэффициентам
разложения бинома Ньютона (а+b)4.
При любом количестве пар генов n частоты
градаций количественного признака в
потомстве будут соответствовать
коэффициентам разложения бинома Ньютона
(а+b)n.
Мы
привели пример, в котором суммируются
эффекты всего двух двухаллельных генов.
Если таких генов будет три, то количество
классов увеличится до 7, а число возможных
комбинаций составит 64, для четырех генов
мы будем иметь 9 классов и 256 комбинаций
и т.д. Простой подсчет показывает, что
при увеличении числа локусов
и аллелей
с суммирующимся эффектом действия на
определенный признак число возможных
генотипов
нарастает очень быстро. Так, если
предположить, что гены какого-либо
количественного признака располагаются
в 20 локусах и имеют по 4 аллеля, то
количество генотипов будет составлять
1020. Понятно, что частотная
гистограмма степени выраженности
признака при таком количестве генотипов
будет все более сглаживаться и напоминать
нормальное распределение. Вся изменчивость
при этом будет носить чисто генетический
характер, и количественная величина,
характеризующая вариативность признака
в популяции (дисперсия), в данном случае
может быть названа генетической
дисперсией.
В рассмотренном примере вся фенотипическая
изменчивость объясняется наследственной
вариативностью, т.е. VP = VG. В
генетике для обозначения дисперсии
используется латинская буква V (от
английского variance). Значками VP VG
и VE обозначаются соответственно
фенотипическая (P = phenotype), генотипическая
(G = genotype) и средовая (Е = environment) дисперсии.
Однако
подобный же непрерывный характер может
носить и изменчивость, целиком определяемая
средовыми факторами. Поэтому, столкнувшись
с непрерывной изменчивостью, невозможно
сразу определить, обусловлена ли она
исключительно воздействиями среды или
здесь имеет место и полигенный
характер наследуемости. В генетике
существуют специальные методы, позволяющие
определить природу изменчивости, но об
этом речь пойдет немного позже. Сейчас
же только подчеркнем, что в большинстве
случаев непрерывный характер изменчивости
определяется как действием многих
генов, так и взаимодействием генотипа
со всевозможными средовыми условиями.
4.2.3. Различные типы взаимодействия генов. Аддитивное взаимодействие
Аддитивное
взаимодействие. Рассмотренный в
предыдущем параграфе пример в некотором
роде является идеальным. В этом идеальном
примере полимерные гены оказывают чисто
аддитивный эффект, то есть происходит
простая суммация действия этих генов.
Предполагается также, что полимерные
гены по силе действия равны друг другу.
Только при соблюдении этих условий
уровень развития количественного
признака в потомстве (Fl, F2) будет строго
промежуточным между родительскими
формами, а кривая распределения генотипов
будет точно соответствовать нормальной.
Чем больше генов бyдет участвовать в
расщеплении, тем меньше в поколении F2
будет доля особей, сходных с исходными
родительскими формами, при этом частоты
градаций количественного признака
будут соответствовать коэффициентам
разложения бинома Ньютона.
Доминирование
и неполное доминирование. Чисто
аддитивный характер взаимодействия
генов встречается достаточно редко,
чаще всего картина является более
сложной. Для некоторых генов могут иметь
место уже известные нам эффекты
доминирования. При этом степень
доминирования может отличаться для
разных генов. В известных нам опытах Г.
Менделя один из аллелей
полностью подавлял другой, то есть имело
место полное доминирование. Однако
могут встречаться и пары аллелей, для
которых доминантность и рецессивность
не проявляются в полной мере. Это случаи
неполного доминирования, или
кодоминантности. В случае неполного
доминирования гетерозиготы
обладают промежуточным фенотипом
между родительскими формами.
Эпистаз.
Взаимодействовать могут не только
аллели одного локуса,
но и аллели, расположенные в разных
локусах. В таких случаях говорят об
эпистатическом взаимодействии, или
просто эпистазе. При эпистазе
присутствие определенного гена (именно
его называют эпистатическим) полностью
подавляет эффект действия другого гена,
расположенного в другом локусе
(подавляемый ген носит название
гипостатического).
Сложные
эффекты взаимодействия генов. Вообще
фенотипические признаки, полностью
контролируемые парой аллелей, расположенных
в одном генном локусе, т.е. подобные т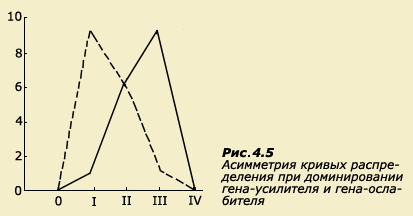 ем,
с которыми имел дело Г. Мендель,
сравнительно редко встречаются в
природе. Большинство признаков
определяется взаимодействием нескольких
генов, находящихся в разных локусах.
При этом эффекты взаимодействия всего
комплекса генов, определяющих данный
фенотипический признак, могут включать
как аддитивный компонент, так и различные
степени доминирования и эпистаз.
Взаимодействующие гены могут как
усиливать эффект действия какого-либо
гена, так и ослаблять его. В результате
кривые распределения генотипов в
потомстве не будут столь идеально
соответствовать кривой нормального
распределения, как при чисто аддитивном
наследовании (рис. 4.5). Распределения
могут оказаться асимметричными и даже
многовершинными.
В
настоящее время принято считать, что в
детерминации количественных признаков
принимают участие различные группы
генов. Существуют главные гены (олигогены),
значительно влияющие на развитие
признака. Существуют гены с более слабым
эффектом. И, наконец, существуют
гены-модификаторы, которые определяют
одни признаки, но одновременно модифицируют
действие главных генов на другие
признаки.
ем,
с которыми имел дело Г. Мендель,
сравнительно редко встречаются в
природе. Большинство признаков
определяется взаимодействием нескольких
генов, находящихся в разных локусах.
При этом эффекты взаимодействия всего
комплекса генов, определяющих данный
фенотипический признак, могут включать
как аддитивный компонент, так и различные
степени доминирования и эпистаз.
Взаимодействующие гены могут как
усиливать эффект действия какого-либо
гена, так и ослаблять его. В результате
кривые распределения генотипов в
потомстве не будут столь идеально
соответствовать кривой нормального
распределения, как при чисто аддитивном
наследовании (рис. 4.5). Распределения
могут оказаться асимметричными и даже
многовершинными.
В
настоящее время принято считать, что в
детерминации количественных признаков
принимают участие различные группы
генов. Существуют главные гены (олигогены),
значительно влияющие на развитие
признака. Существуют гены с более слабым
эффектом. И, наконец, существуют
гены-модификаторы, которые определяют
одни признаки, но одновременно модифицируют
действие главных генов на другие
признаки.
4.2.4. Возникновение количественной изменчивости под действием среды. Норма реакции. Средовая дисперсия
В двух предыдущих
разделах мы позволили себе на время
забыть о влиянии среды на выраженность
количественного признака и рассмотрели
пример возникновения количественной
изменчивости только под влиянием
совместного действия генов на один и
тот же признак. Сейчас совершим обратную
операцию - представим себе, что в популяции
отсутствует генетическая изменчивость.
Что это означает? Только то, что вся
популяция состоит из особей с одинаковыми
генотипами, т.е. никакого генетического
разнообразия нет, а все наблюдаемое
разнообразие фенотипов определяется
только тем, что каждая особь развивалась
в своих, отличных от других, условиях
среды. Такие генетические однородные
популяции (или клоны), в принципе,
можно получить у простейших организмов
или растений, размножающихся вегетативным
делением. Существуют также и специально
выведенные линии лабораторных животных,
обладающих одинаковым генотипом по
какому-либо признаку - так называемые
чистые линии, получаемые путем
близкородственного скрещивания (такой
тип скрещивания называется инбридингом,
а получаемые при этом чистые линии
животных называются инбредными).
Таким образом, когда мы говорим о
популяции организмов с одинаковыми
генотипами, мы не оказываемся в области
чисто теоретических рассуждений, но
можем опираться на данные реальных
экспериментов.
Итак,
для некоторых видов живых организмов
мы можем получить большое число особей
с одинаковыми генотипами. Мы можем
поместить их в различные фиксированные
условия среды. Например, выращивать при
различной температуре, при различном
содержании кислорода или каких-либо
питательных веществ и т.д. В результате
мы получим множество особей, все различия
между которыми будут обусловлены только
разницей в средовых условиях. Многие
знакомы с вегетативным размножением
растений. Когда мы высаживаем на грядку
"усы" от одного растения земляники,
мы получаем клон генетически идентичных
кустиков. Все различия между ними (в
плодовитости, размерах ягод, листьев и
т.д.) будут обусловлены не наследственными
причинами, поскольку генотипы всех
растений одинаковы, а различиями в
условиях выращивания (состав почвы,
освещение, количество влаги и
т.п.).
Итак,
допустим, что генотип представляет
собой константу, и рассмотрим фенотип
к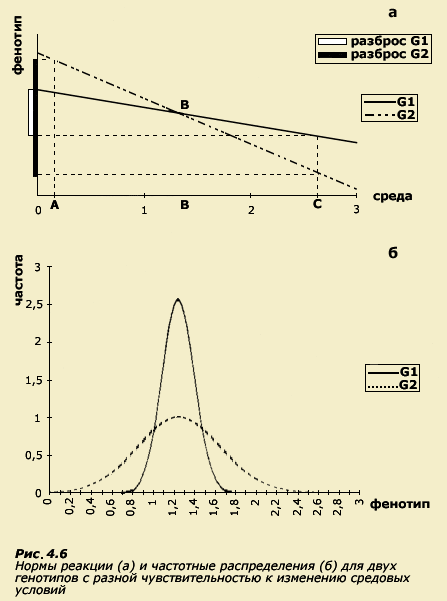 ак
функцию условий внешней среды. Для этого
нам необходимо ввести еще одно важное
понятие генетики - это понятие нормы
реакции. Если данный генотип чувствителен
к среде, то, поместив множество одинаковых
генотипов в различные строго фиксированные
условия среды, мы получим множество
отличающихся фенотипов. Мы можем измерить
выраженность интересующего нас признака
и построить график зависимости
характеристик фенотипа от параметров
среды. Экспериментально это возможно
осуществить для клонов или чистых линий
различных организмов. Полученные в
результате графики зависимости
особенностей фенотипа от параметров
среды и будут отображать нормы реакции.
Каждый генотип будет характеризоваться
своей зависимостью, т.е. своей нормой
реакции.
Рассмотрим
гипотетический пример. Предположим, мы
имеем две инбредные линии мышей с
генотипами G1 и G2. Нас, например, интересует
зависимость густоты шерсти (фенотип)
от температурных условий (среда), в
которых выращиваются эти мыши. Допустим,
эта зависимость носит линейный характер:
чем выше температура, тем меньше густота
шерсти животных. Но генотипы могут
отличаться своей чувствительностью к
температурным условиям. Предположим,
что G1 оказался менее чувствительным к
температуре, чем G2. Соответственно, если
мы построим графики зависимости фенотипа
от среды (нормы реакции), то для генотипа
G1 будет характерен меньший наклон прямой
(рис. 4.6). На нашем рисунке существует
точка пересечения, в которой при
определенной температуре фенотипы
животных, принадлежащих к разным
инбредным линиям, будут совпадать.
Зададим теперь такой вопрос: различаются
ли фенотипически (по густоте шерсти)
животные обеих линий? Понятно, что
однозначного ответа на этот вопрос дать
нельзя, поскольку существуют такие
условия среды (точка В), в которых фенотипы
не различаются. Нельзя также ответить
и на вопрос, животные какой линии имеют
более густую шерсть, поскольку в одних
температурных условиях более густую
шерсть имеют животные с генотипом G1, а
в других - с генотипом G2. Единственное,
в чем мы можем быть уверены, так это в
том, что генотипы различаются своими
нормами реакции.
Проделаем
теперь такую процедуру. Из точек А и С,
находящихся на полюсах оси средовых
изменений, проведем проекции на графики
норм реакции обоих генотипов, а затем
из точек пересечения опустим проекции
на ось фенотипов. Мы увидим, что в одном
и том же средовом диапазоне вариативность
фенотипов (разброс) более чувствительного
к среде генотипа G2 окажется выше, чем
для менее чувствительного генотипа G1.
Если отобразить это на графике
распределений (рис. 4.6 б), мы увидим, что
генотипы различаются по разбросу
значений вокруг среднего, то есть они
отличаются своими дисперсиями: для
генотипа G2 характерна более высокая
дисперсия, чем для генотипа G1. Дисперсия
величины густоты шерсти внутри каждой
группы мышей обусловлена чисто средовыми
причинами и может быть названа средовой
дисперсией. Различия в дисперсиях
между разными линиями объясняются
разной чувствительностью генотипов к
среде: чем выше чувствительность данного
генотипа к среде, тем выше фенотипическая
дисперсия, порождаемая
особенностями среды.
Таким
образом, можно сказать, что генотип
однозначно не определяет фенотип.
Чтобы правильно описать сравниваемые
генотипы, недостаточно сказать, что они
определяют разные фенотипы, поскольку
при определенных средовых условиях это
утверждение может оказаться ложным.
Лучше сравнивать генотипы по характерным
для них нормам реакции. Кроме того,
следует подчеркнуть, что генотипы
отличаются по их чувствительности к
средовым воздействиям. Более
чувствительному генотипу соответствует
и большая средовая дисперсия.
Существуют
примеры нормы реакции, при которой
количественные изменения в окружающей
среде могут привести даже к качественному
изменению фенотипа, несмотря на то, что
генотип остается одним и тем же. Таким
примером может служить изменение окраски
цветков Primula sinensis. Это растение в
обычных условиях имеет красную окраску
цветков. Когда то же самое растение
переносят в оранжерею, где поддерживается
температура 30-35°, то через некоторое
время вновь расцветающие цветки
приобретают белую окраску, т.е. происходит
качественное изменение фенотипа в ответ
на изменение среды.
Таким
образом, можно сказать, что норма
реакции - это специфический
характер реакции данного генотипа на
изменение окружающих условий. Генетики
говорят, что наследуются не определенные
признаки, а нормы реакций.
Существует
еще одно близкое по смыслу понятие - это
понятие диапазона реакции. Под
диапазоном реакции в генетике
обычно понимают размах значений фенотипа
в определенных границах среды. В нашем
вымышленном примере диапазон изменений
густоты шерсти в определенных границах
температурных условий и будет являться
диапазоном реакции.
Приведенные
примеры показывают, что для животных и
растений, если мы располагаем достаточным
количеством особей с одинаковыми
генотипами, мы имеем реальную возможность
изучить реальные нормы реакции на
изменение средовых условий. Сделать то
же самое в отношении человека практически
невозможно. Невозможность получить
нормы реакции для признаков человека
объясняется двумя причинами. Во-первых,
у человека нельзя получить большое
число генетически идентичных организмов,
чтобы затем выращивать их в различных
условиях. Максимальное количество
генетически идентичных людей соответствует
тем клонам монозиготных близнецов,
которые доступны для изучения (как
правило, не более пяти человек). Во-вторых,
недопустимо подвергать людей различным
средовым воздействиям в экспериментальных
целях. Кроме того, для большинства
психологических признаков пока даже
не известно, какие особенности среды
следует измерять.
Таким
образом, точные нормы реакции для
различных признаков человека, и тем
более для психологических характеристик,
получить практически невозможно. Однако
на основании некоторых косвенных данных
возможно гипотетическое моделирование
нормы реакции.
В
психологии для времени сенсомоторной
р
ак
функцию условий внешней среды. Для этого
нам необходимо ввести еще одно важное
понятие генетики - это понятие нормы
реакции. Если данный генотип чувствителен
к среде, то, поместив множество одинаковых
генотипов в различные строго фиксированные
условия среды, мы получим множество
отличающихся фенотипов. Мы можем измерить
выраженность интересующего нас признака
и построить график зависимости
характеристик фенотипа от параметров
среды. Экспериментально это возможно
осуществить для клонов или чистых линий
различных организмов. Полученные в
результате графики зависимости
особенностей фенотипа от параметров
среды и будут отображать нормы реакции.
Каждый генотип будет характеризоваться
своей зависимостью, т.е. своей нормой
реакции.
Рассмотрим
гипотетический пример. Предположим, мы
имеем две инбредные линии мышей с
генотипами G1 и G2. Нас, например, интересует
зависимость густоты шерсти (фенотип)
от температурных условий (среда), в
которых выращиваются эти мыши. Допустим,
эта зависимость носит линейный характер:
чем выше температура, тем меньше густота
шерсти животных. Но генотипы могут
отличаться своей чувствительностью к
температурным условиям. Предположим,
что G1 оказался менее чувствительным к
температуре, чем G2. Соответственно, если
мы построим графики зависимости фенотипа
от среды (нормы реакции), то для генотипа
G1 будет характерен меньший наклон прямой
(рис. 4.6). На нашем рисунке существует
точка пересечения, в которой при
определенной температуре фенотипы
животных, принадлежащих к разным
инбредным линиям, будут совпадать.
Зададим теперь такой вопрос: различаются
ли фенотипически (по густоте шерсти)
животные обеих линий? Понятно, что
однозначного ответа на этот вопрос дать
нельзя, поскольку существуют такие
условия среды (точка В), в которых фенотипы
не различаются. Нельзя также ответить
и на вопрос, животные какой линии имеют
более густую шерсть, поскольку в одних
температурных условиях более густую
шерсть имеют животные с генотипом G1, а
в других - с генотипом G2. Единственное,
в чем мы можем быть уверены, так это в
том, что генотипы различаются своими
нормами реакции.
Проделаем
теперь такую процедуру. Из точек А и С,
находящихся на полюсах оси средовых
изменений, проведем проекции на графики
норм реакции обоих генотипов, а затем
из точек пересечения опустим проекции
на ось фенотипов. Мы увидим, что в одном
и том же средовом диапазоне вариативность
фенотипов (разброс) более чувствительного
к среде генотипа G2 окажется выше, чем
для менее чувствительного генотипа G1.
Если отобразить это на графике
распределений (рис. 4.6 б), мы увидим, что
генотипы различаются по разбросу
значений вокруг среднего, то есть они
отличаются своими дисперсиями: для
генотипа G2 характерна более высокая
дисперсия, чем для генотипа G1. Дисперсия
величины густоты шерсти внутри каждой
группы мышей обусловлена чисто средовыми
причинами и может быть названа средовой
дисперсией. Различия в дисперсиях
между разными линиями объясняются
разной чувствительностью генотипов к
среде: чем выше чувствительность данного
генотипа к среде, тем выше фенотипическая
дисперсия, порождаемая
особенностями среды.
Таким
образом, можно сказать, что генотип
однозначно не определяет фенотип.
Чтобы правильно описать сравниваемые
генотипы, недостаточно сказать, что они
определяют разные фенотипы, поскольку
при определенных средовых условиях это
утверждение может оказаться ложным.
Лучше сравнивать генотипы по характерным
для них нормам реакции. Кроме того,
следует подчеркнуть, что генотипы
отличаются по их чувствительности к
средовым воздействиям. Более
чувствительному генотипу соответствует
и большая средовая дисперсия.
Существуют
примеры нормы реакции, при которой
количественные изменения в окружающей
среде могут привести даже к качественному
изменению фенотипа, несмотря на то, что
генотип остается одним и тем же. Таким
примером может служить изменение окраски
цветков Primula sinensis. Это растение в
обычных условиях имеет красную окраску
цветков. Когда то же самое растение
переносят в оранжерею, где поддерживается
температура 30-35°, то через некоторое
время вновь расцветающие цветки
приобретают белую окраску, т.е. происходит
качественное изменение фенотипа в ответ
на изменение среды.
Таким
образом, можно сказать, что норма
реакции - это специфический
характер реакции данного генотипа на
изменение окружающих условий. Генетики
говорят, что наследуются не определенные
признаки, а нормы реакций.
Существует
еще одно близкое по смыслу понятие - это
понятие диапазона реакции. Под
диапазоном реакции в генетике
обычно понимают размах значений фенотипа
в определенных границах среды. В нашем
вымышленном примере диапазон изменений
густоты шерсти в определенных границах
температурных условий и будет являться
диапазоном реакции.
Приведенные
примеры показывают, что для животных и
растений, если мы располагаем достаточным
количеством особей с одинаковыми
генотипами, мы имеем реальную возможность
изучить реальные нормы реакции на
изменение средовых условий. Сделать то
же самое в отношении человека практически
невозможно. Невозможность получить
нормы реакции для признаков человека
объясняется двумя причинами. Во-первых,
у человека нельзя получить большое
число генетически идентичных организмов,
чтобы затем выращивать их в различных
условиях. Максимальное количество
генетически идентичных людей соответствует
тем клонам монозиготных близнецов,
которые доступны для изучения (как
правило, не более пяти человек). Во-вторых,
недопустимо подвергать людей различным
средовым воздействиям в экспериментальных
целях. Кроме того, для большинства
психологических признаков пока даже
не известно, какие особенности среды
следует измерять.
Таким
образом, точные нормы реакции для
различных признаков человека, и тем
более для психологических характеристик,
получить практически невозможно. Однако
на основании некоторых косвенных данных
возможно гипотетическое моделирование
нормы реакции.
В
психологии для времени сенсомоторной
р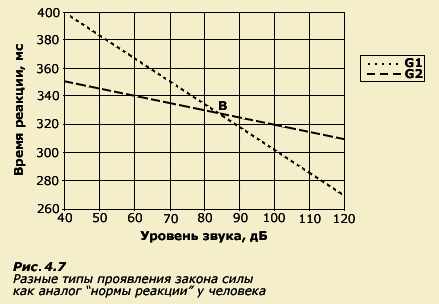 еакции
человека известно такое явление, как
закон силы. Состоит оно в следующем.
Если человека просят как можно быстрее
нажимать на кнопку в ответ на звуковой
сигнал и при этом регистрируют точное
время между началом сигнала и моментом
нажатия, то в результате получают
количественную характеристику (измеряемую
в миллисекундах), которая называется
временем сенсомоторной реакции (ВСМР).
Можно считать ВСМР количественным
фенотипическим признаком человека, тем
более что все люди реагируют с различной
скоростью. Если интенсивность сигнала
менять от слабой к сильной, то ВСМР будет
сокращаться. Вот эту зависимость в
психологии и называют законом
силы.
Оказалось,
что закон силы у разных людей проявляется
по-разному. Одни люди дают резкое
сокращение ВСМР, а график ее зависимости
от интенсивности стимула характеризуется
крутым наклоном. Для графиков других
людей характерно более пологое снижение.
В работах известного исследователя
индивидуальных различий В.Д.
Небылицына было показано, что
различия в характере действия интенсивности
стимула связаны со свойствами нервной
системы.
Предположим,
что в популяции существуют крайние
варианты генотипов, для которых характерны
разные типы реакции на изменение
интенсивности звука. Можно эти зависимости
рассматривать как аналог нормы реакции,
где фенотип
- это ВСМР, а среда - это разная интенсивность
звука. Построим графики "нормы реакции"
для двух гипотетических генотипов,
отличающихся характером закона силы.
На рисунке 4.7 мы изобразили эти зависимости.
Гипотетический генотип G1 характеризуется
крутым наклоном графика зависимости
ВСМР от интенсивности звука, а
гипотетический генотип G2 - пологим
наклоном. Так же как и в случае с мышами,
мы видим, что существует среда (точка
В), в пределах которой генотипы
фенотипически не различаются. Левее
этой точки генотип G2 опережает другой
генотип (G1) по скорости реакции, а правее
- наоборот, более высокая скорость
реагирования характерна для генотипа
G1. Таким образом, не существует абсолютных
различий в фенотипах, а все зависит от
диапазона среды, в котором проводятся
измерения.
Рассмотренное
в этом разделе понятие нормы реакции
представляет определенный практический
интерес для психолога и педагога.
Поскольку каждый человек обладает
уникальным генотипом, он обладает и
уникальной нормой реакции на те или
иные средовые воздействия, то есть
каждый генотип по-своему реагирует на
одни и те же условия среды. Следовательно,
поместив группу детей в унифицированные
условия среды (примером может служить
обучение в одном классе), мы не вправе
ожидать одинакового результата от всех
детей - каждый будет реагировать по-своему
и характеризоваться своим уровнем
достижений в соответствии с нормой
реакции своего генотипа. Каждый из нас
генетически уникален, и с этим необходимо
считаться в практике обучения и
воспитания, создавая максимальное
разнообразие средовых условий развития.
еакции
человека известно такое явление, как
закон силы. Состоит оно в следующем.
Если человека просят как можно быстрее
нажимать на кнопку в ответ на звуковой
сигнал и при этом регистрируют точное
время между началом сигнала и моментом
нажатия, то в результате получают
количественную характеристику (измеряемую
в миллисекундах), которая называется
временем сенсомоторной реакции (ВСМР).
Можно считать ВСМР количественным
фенотипическим признаком человека, тем
более что все люди реагируют с различной
скоростью. Если интенсивность сигнала
менять от слабой к сильной, то ВСМР будет
сокращаться. Вот эту зависимость в
психологии и называют законом
силы.
Оказалось,
что закон силы у разных людей проявляется
по-разному. Одни люди дают резкое
сокращение ВСМР, а график ее зависимости
от интенсивности стимула характеризуется
крутым наклоном. Для графиков других
людей характерно более пологое снижение.
В работах известного исследователя
индивидуальных различий В.Д.
Небылицына было показано, что
различия в характере действия интенсивности
стимула связаны со свойствами нервной
системы.
Предположим,
что в популяции существуют крайние
варианты генотипов, для которых характерны
разные типы реакции на изменение
интенсивности звука. Можно эти зависимости
рассматривать как аналог нормы реакции,
где фенотип
- это ВСМР, а среда - это разная интенсивность
звука. Построим графики "нормы реакции"
для двух гипотетических генотипов,
отличающихся характером закона силы.
На рисунке 4.7 мы изобразили эти зависимости.
Гипотетический генотип G1 характеризуется
крутым наклоном графика зависимости
ВСМР от интенсивности звука, а
гипотетический генотип G2 - пологим
наклоном. Так же как и в случае с мышами,
мы видим, что существует среда (точка
В), в пределах которой генотипы
фенотипически не различаются. Левее
этой точки генотип G2 опережает другой
генотип (G1) по скорости реакции, а правее
- наоборот, более высокая скорость
реагирования характерна для генотипа
G1. Таким образом, не существует абсолютных
различий в фенотипах, а все зависит от
диапазона среды, в котором проводятся
измерения.
Рассмотренное
в этом разделе понятие нормы реакции
представляет определенный практический
интерес для психолога и педагога.
Поскольку каждый человек обладает
уникальным генотипом, он обладает и
уникальной нормой реакции на те или
иные средовые воздействия, то есть
каждый генотип по-своему реагирует на
одни и те же условия среды. Следовательно,
поместив группу детей в унифицированные
условия среды (примером может служить
обучение в одном классе), мы не вправе
ожидать одинакового результата от всех
детей - каждый будет реагировать по-своему
и характеризоваться своим уровнем
достижений в соответствии с нормой
реакции своего генотипа. Каждый из нас
генетически уникален, и с этим необходимо
считаться в практике обучения и
воспитания, создавая максимальное
разнообразие средовых условий развития.
4.2.5. Совместный вклад генотипа и среды в количественную изменчивость. Генетическая и средовая дисперсии как составляющие популяционной фенотипической дисперсии
Фенотипические различия между людьми объясняются по крайней мере двумя причинами. Во-первых, люди отличаются друг от друга своими генотипами. Это приводит к возникновению генетически обусловленной изменчивости. Во-вторых, каждый человек развивается в особенных средовых условиях. Это приводит к возникновению средовой изменчивости. Теперь опишем, как общая популяционная изменчивость признаков складывается из генетической и средовой изменчивости. Рассмотрим компьютерную модель количественной изменчивости в популяции (автором модели является Л.С. Куравский). Допустим, мы измерили некий количественный признак у 1000 индивидов в какой-либо популяции и построили соответствующее частотное распределение. Будем считать, что наш признак определяется одним геном, который может существовать в трех различных формах, т.е. имеет три аллеля. Обозначим их буквами А, В и С. Тогда в популяции могут встретиться три типа гомозигот - АА, ВВ, СС - и три типа гетерозигот - АВ, ВС и АС. Пусть люди с генотипом АА имеют самые низкие значения признака, а люди с генотипом СС - самые высокие. Генотип ВВ занимает промежуточное положение. Эффекты доминирования отсутствуют, и гетерозиготы оказываются промежуточными между гомозиготами. Мы знаем, что на конкретную величину признака оказывают влияние не только гены, но и средовые условия, например, характер питания или родительской заботы в детстве. Распределение фенотипов для каждого генотипа будет характеризоваться определенным средним значением и разбросом вокруг среднего, или дисперсией. Величина дисперсии будет определяться тем, насколько данный генотип чувствителен к средовым влияниям. Чем выше чувствительность генотипа к среде, тем большее разнообразие фенотипов мы получим и соответственно тем большей дисперсией будет характеризоваться данный генотип (см. предыдущий параграф). Понятно, что внутри группы особей с одинаковым генотипом дисперсия будет определяться только средовыми факторами и поэтому будет чисто средовой. Например, для представителей генотипа АА: VP(AA) = VE(AA) Предположим, мы смогли разделить людей, обладающих конкретными генотипами, на соответствующие группы, то есть мы разбили всю популяцию на шесть групп в соответствии с генотипом каждого человека. Обычно отдельные генотипы представлены в популяции неравномерно, поэтому можно ожидать, что и обладатели наших вымышленных генотипов встречаются с определенной частотой.
Таким образом, мы можем измерить:
среднее значение признака для каждого генотипа,
частоту его встречаемости в популяции,
величину разброса индивидуальных значений вокруг среднего, то есть дисперсию VE(AA), VE(BB) и т.д.
Представим себе,
что мы провели все измерения, получили
статистические оценки и занесли их в
таблицу (количественные значения
признака были выбраны таким образом,
чтобы они примерно соответствовали
реальному количественному признаку
человека, каковым является рост в
сантиметрах, поэтому можно представить
себе, что мы имеем дело с формированием
популяционной изменчивости по росту,
хотя, конечно же, генетическая детерминация
р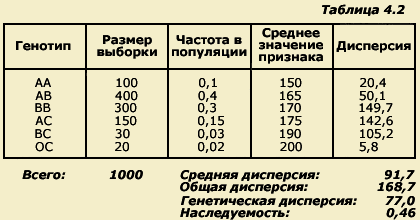 еального
роста отличается от схемы, приведенной
в данной модели). Одновременно мы
построили частотные распределения для
каждого конкретного генотипа и
общепопуляционное распределение (рис.
4.8).
На
графике мы видим шесть субраспределений,
каждое из которых описывает один из
генотипов в популяции. Распределение
каждого генотипа представлено
пропорционально его встречаемости в
популяции. Наибольшей представленностью
на графике характеризуется генотип АВ,
п
еального
роста отличается от схемы, приведенной
в данной модели). Одновременно мы
построили частотные распределения для
каждого конкретного генотипа и
общепопуляционное распределение (рис.
4.8).
На
графике мы видим шесть субраспределений,
каждое из которых описывает один из
генотипов в популяции. Распределение
каждого генотипа представлено
пропорционально его встречаемости в
популяции. Наибольшей представленностью
на графике характеризуется генотип АВ,
п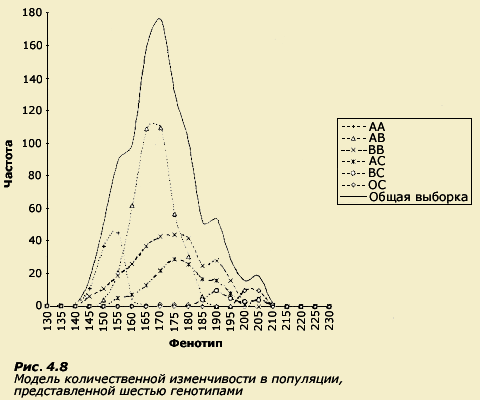 оскольку
этот генотип более всего распространен
в популяции, а наименьшей - СС. Одни
генотипы (АА, АВ) характеризуются
небольшими средними значениями признака,
другие (ВС, СС) - большими. Каждому из
генотипов соответствует определенное
разнообразие фенотипов (дисперсия),
поскольку обладатели этого генотипа
росли в разных средах. Если генотип
очень чувствителен к среде (например,
BB и АС), ему будет соответствовать большая
фенотипическая дисперсия, а если
малочувствителен (например, СС), то
выраженность признака у всех обладателей
этого генотипа будет близка к генотипической
средней. Форма же общепопуляционного
распределения является следствием
сложения всех частных распределений
для каждого из генотипов.
Теперь
обратимся к таблице 4.2. В одной из колонок
выписаны значения средних величин
признака для каждого генотипа. Наибольший
интерес представляет последняя колонка,
в которой представлены дисперсии. Мы
видим, что дисперсии генотипов значительно
различаются вследствие их разной
чувствительности к средовым влияниям.
При этом дисперсия внутри каждого
генотипа целиком определяется влияниями
среды. Общая же изменчивость в популяции
и характеризующая ее величина
общепопуляционной дисперсии складывается
из двух составляющих. Одна из них - это
различия между генотипами, выражающиеся
в различиях между средними значениями
признака у обладателей отдельных
генотипов (АА, АВ и т.д.), а другая - это
различия внутри каждого из генотипов,
которые определяются только средой. В
нашем примере генетическая составляющая
дисперсии возникает при участии только
шести генотипов, то есть генетическая
дисперсия возникла в результате различий
между средними значениями признака
имеющихся у нас шести генотипов. Если
на графике убрать кривые распределений,
а оставить только столбцы, соответствующие
частоте встречаемости средних значений
признака этих шести генотипов, то мы
будем иметь шесть таких столбцов,
представляющих гистограмму распределения
средних значений признака всех генотипов
нашей популяции. В реальных популяциях,
где имеется множество различных
генотипов, такая гистограмма будет
состоять не из шести столбцов, а из сотен
и тысяч, а распределение будет
соответствовать нормальному. Средовая
составляющая общепопуляционной дисперсии
складывается из отдельных средовых
дисперсий внутри каждого генотипа.
Таким образом, общепопуляционная
дисперсия состоит из средовой дисперсии
внутри отдельных генотипов и генетической
дисперсии, возникающей за счет различий
между генотипами, что можно выразить
формулой
оскольку
этот генотип более всего распространен
в популяции, а наименьшей - СС. Одни
генотипы (АА, АВ) характеризуются
небольшими средними значениями признака,
другие (ВС, СС) - большими. Каждому из
генотипов соответствует определенное
разнообразие фенотипов (дисперсия),
поскольку обладатели этого генотипа
росли в разных средах. Если генотип
очень чувствителен к среде (например,
BB и АС), ему будет соответствовать большая
фенотипическая дисперсия, а если
малочувствителен (например, СС), то
выраженность признака у всех обладателей
этого генотипа будет близка к генотипической
средней. Форма же общепопуляционного
распределения является следствием
сложения всех частных распределений
для каждого из генотипов.
Теперь
обратимся к таблице 4.2. В одной из колонок
выписаны значения средних величин
признака для каждого генотипа. Наибольший
интерес представляет последняя колонка,
в которой представлены дисперсии. Мы
видим, что дисперсии генотипов значительно
различаются вследствие их разной
чувствительности к средовым влияниям.
При этом дисперсия внутри каждого
генотипа целиком определяется влияниями
среды. Общая же изменчивость в популяции
и характеризующая ее величина
общепопуляционной дисперсии складывается
из двух составляющих. Одна из них - это
различия между генотипами, выражающиеся
в различиях между средними значениями
признака у обладателей отдельных
генотипов (АА, АВ и т.д.), а другая - это
различия внутри каждого из генотипов,
которые определяются только средой. В
нашем примере генетическая составляющая
дисперсии возникает при участии только
шести генотипов, то есть генетическая
дисперсия возникла в результате различий
между средними значениями признака
имеющихся у нас шести генотипов. Если
на графике убрать кривые распределений,
а оставить только столбцы, соответствующие
частоте встречаемости средних значений
признака этих шести генотипов, то мы
будем иметь шесть таких столбцов,
представляющих гистограмму распределения
средних значений признака всех генотипов
нашей популяции. В реальных популяциях,
где имеется множество различных
генотипов, такая гистограмма будет
состоять не из шести столбцов, а из сотен
и тысяч, а распределение будет
соответствовать нормальному. Средовая
составляющая общепопуляционной дисперсии
складывается из отдельных средовых
дисперсий внутри каждого генотипа.
Таким образом, общепопуляционная
дисперсия состоит из средовой дисперсии
внутри отдельных генотипов и генетической
дисперсии, возникающей за счет различий
между генотипами, что можно выразить
формулой
VP = VG + VE.
В рассмотренном примере количественная оценка общей дисперсии оказалась равной 168,7, а усредненная оценка средовой дисперсии - 91,7, то есть почти вполовину меньше. Если из величины общей дисперсии вычесть величину средовой дисперсии, мы получим оценку генетической дисперсии. Таким образом, в нашем примере генетическая дисперсия равна:
168,7-91,7=77,0.
Это означает, что генетическая дисперсия обусловливает чуть меньше половины всех индивидуальных различий в популяции.
Выводы
Вся совокупность генов организма составляет его генотип.
Любые проявления организма в каждый момент жизни составляют его фенотип.
Фенотип есть результат взаимодействия генотипа со средой.
Количественная изменчивость может возникать в результате полимерного действия многих генов на один признак.
Дисперсия количественного признака, возникающая за счет действия генов, носит название генетической дисперсии.
Существуют различные формы взаимодействия генов: аддитивное, доминирование, эпистаз и др.
Количественная изменчивость может возникать под действием факторов среды.
Дисперсия количественного признака, возникающая за счет средовых влияний, носит название средовой дисперсии.
Генотипы по-разному реагируют на одни и те же изменения среды. Специфический характер реакции данного reнотипа на изменение окружающих условий носит название нормы реакции.
Генотипы отличаются по своей чувствительности к среде.
Фенотипическая изменчивость в популяции складывается из генетической и средовой изменчивости; фенотипическая популяционная дисперсия признака представляет собой сумму его генетической и средовой дисперсий.
4.3. Показатель наследуемости и его особенности
4.3.1. Показатель наследуемости в количественной генетике и генетике поведения
4.3.2. Чувствительность показателя наследуемости к частотам генотипов в популяции
4.3.3. Чувствительность показателя наследуемости к изменениям среды
4.3.4. Важность правильной интерпретации показателя наследуемости
4.3.1. Показатель наследуемости в количественной генетике и генетике поведения
В предыдущем разделе мы продемонстрировали, как из генетической и средовой изменчивости признака складывается его фенотипическая изменчивость. Как было показано на примере, основными компонентами фенотипической дисперсии признака являются генетическая и средовая дисперсии. В количественной генетике долю генетической составляющей в общей фенотипической дисперсии признака принято называть наследуемостью в широком смысле слова, или просто наследуемостью. Наследуемость обычно обозначают символом h2. Наследуемость можно подсчитать, если величину генетической дисперсии разделить на величину общей дисперсии:
 или
или
![]()
Если использовать данные рассмотренного нами примера, то
![]()
Таким образом, доля генетической составляющей в фенотипической дисперсии в нашем примере составляет 46%. Это означает, что изменчивость (вариативность) нашего признака в популяции на 46% складывается за счет вариативности генотипов и на 54% - за счет вариативности сред. Показатель наследуемости - это генетическая составляющая дисперсии. Поэтому при символе наследуемости всегда стоит значок квадрата, указывающий на то, что показатель наследуемости есть компонент дисперсии. Наследуемость является одним из основных показателей, которыми оперирует генетика поведения. Если наследуемость отлична от нуля, это означает, что в основе изменчивости признака лежат не только средовые, но и генетические причины. Когда задается вопрос, лежат ли в основе индивидуальных психологических различий наследственные причины, для ответа необходимо получить данные о влиянии генов на фенотипическую изменчивость. Величина коэффициента наследуемости и позволяет оценить меру влияния генотипа на изменчивость признака. Вся история генетики поведения до некоторого времени представляла собой бесконечные попытки обосновать существование наследственных причин межиндивидуальных различий в поведении. После создания теории наследственности количественных признаков и разработки математического аппарата для анализа генетических и средовых дисперсий практически в каждой работе по генетике поведения человека приводятся данные о наследуемости изучавшейся психологической характеристики. Когда в печати стали появляться сообщения о том, что наследуемость умственных способностей составляет 70-80%, это привлекло внимание не только ученых, но и общества в целом, и оказало в некоторых странах влияние на социальную политику в отношении умственно неполноценных людей. Если коэффициент интеллекта имеет высокую наследуемость, стоит ли тратить силы и средства на специальное обучение индивидов со сниженным интеллектом? И в целом ряде случаев из-за некомпетентного подхода к пониманию наследуемости общество предпринимало ошибочные шаги в решении отдельных социальных проблем. (Об этом более подробно уже говорилось в теме 1). Негативные социальные последствия психогенетических исследований интеллекта явились в том числе результатом очень распространенного заблуждения, связанного с неверным толкованием понятия "наследуемость". К сожалению, очень часто научное понятие наследуемости смешивается с обыденным пониманием наследуемого как чего-то рокового, не подверженного изменению под влиянием средовых условий. Возможно, это обусловлено самой терминологией, поскольку слово "наследуемость" сходно по звучанию со словом "наследственность". Остановимся более подробно на том, какое же содержание вкладывается в научное понятие наследуемости.
4.3.2. Чувствительность показателя наследуемости к частотам генотипов в популяции
Итак, мы выяснили, что наследуемость, вернее, показатель, или коэффициент наследуемости, является генетической компонентой фенотипической дисперсии в популяции. Поскольку популяция не является стабильным образованием, в ней могут происходить процессы, приводящие к изменению состава генотипов. Посмотрим, скажется ли это на показателе наследуемости. Обратимся опять к описанной выше популяционной модели, в которой количественная изменчивость складывается из вариативности признака у обладателей шести генотипов. Предположим, что по каким-либо причинам из популяции исчез аллель А. Если такое произойдет, то наша популяция будет представлена всего тремя генотипами - ВВ, ВС и СС. Соответственно изменятся и количественные величины дисперсий - общепопуляционной, генетической и средовой. А поскольку наследуемость представляет собой отношение генетической дисперсии к общей дисперсии признака, такие изменения в составе генотипов неминуемо отразятся и на показателе наследуемости. Отсюда следует, что наследуемость не является атрибутом признака как такового, а зависит от состава генотипов той популяции, на которой проведено исследование. В другой популяции с другим составом генотипов наследуемость того же самого признака может оказаться иной. Наследуемость может измениться и в том случае, когда один и тот же признак будет изучаться на той же территории, но спустя какой-то срок, поскольку за определенный период состав генотипов в популяции может измениться вследствие миграции или других причин. Если допустить, что в нашей гипотетической популяции исчезнет не один аллель, а сразу два, например, А и С, то популяция будет представлена только одним генотипом - гомозиготами ВВ, а это означает, что вся дисперсия окажется только средовой, и коэффициент наследуемости будет равен нулю, поскольку никакой генетической дисперсии в популяции не будет. Пример такой крайней ситуации особенно отчетливо показывает, что показатель наследуемости есть популяционная характеристика, чувствительная к изменению частот различных генотипов в популяции.
4.3.3. Чувствительность показателя наследуемости к изменениям среды
Мы уже видели, что
показатель наследуемости зависит от
состава генотипов. Если в популяции
меняются частоты генотипов, это приводит
и к изменению показателя наследуемости.
Но в популяции могут происходить и
другие процессы, связанные со сменой
средовых условий. Изменения в среде
также могут отражаться на изменчивости.
Например, при смене времен года хорошо
заметны изменения в цвете кожи человека.
Зимой люди с бледной и смуглой кожей
отчетливо различаются. Летом под
действием солнца кожа темнеет и различия
между людьми по цвету кожи становятся
менее заметными, следовательно,
изменчивость уменьшается. Рассмотрим,
может ли изменение среды отразиться на
показателе наследуемости. Для удобства
воспользуемся уже описанной моделью
нормы реакции на примере закона силы
(рис. 4.7 и 4.9).
Предположим,
что популяция представлена двумя
генотипами (G1 и G2). Каждый генотип
характеризуется своей нормой реакции,
т.е. специфическим характером реагирования
на изменение окружающих условий. В нашем
случае это время сенсомоторной
реакции (ВСМР) на звуки
разной интенсивности. На рисунке 4.9а
изображены графики нормы реакции каждого
генотипа. На оси ординат отложены
значения фенотипа - в нашем примере это
величины ВСМР, а в принципе это может
быть любая количественная величина,
поддающаяся измерению. На оси абсцисс
отложены количественные значения
параметров среды. В качестве средового
параметра у нас выступает интенсивность
звука, но теоретически это может быть
любая характеристика среды (температура,
концентрация какого-либо вещества в
субстрате и иные поддающиеся контролю
величины). Мы видим, что генотип G1 более
чувствителен к среде, чем генотип G2.
Графики норм реакции обоих генотипов
пересекаются. Теперь рассмотрим, какие
фенотипы будут представлены в популяции
в различных диапазонах сред. На графике
эти диапазоны обозначены тремя отрезками
(среда 1, среда 2 и среда 3). Спроецируем
эти отрезки на сами графики, и затем из
точек пересечения проведем перпендикуляры
на ось ординат, чтобы получить крайние
значения фенотипов для генотипов G1 и
G2 в каждом средовом диапазоне. На рисунке
4.9 б, в, г мы изобразили, какие фенотипы
мы встретим в популяции в указанных
диапазонах среды: на графиках представлены
частотные распределения по каждому
генотипу и по всей популяции в целом в
средовых диапазонах 1, 2 и 3. Мы видим, что
в диапазоне 2 (рис. 4.9 в) средние величины
признака для обоих генотипов совпадают.
Это означает, что здесь отсутствует
дисперсия, возникающая за счет различий
в средних значениях признака соответствующих
генотипов, а мы знаем, что это и есть
генетическая дисперсия. Значит, в нашем
случае вся дисперсия представлена
только средовой составляющей. Единственное
отличие заключается в большей
чувствительности генотипа G1 к среде. В
результате распределение по популяции
в целом будет выглядеть так, как показано
на рисунке. Здесь одна мода,
и практически нет генетической дисперсии
(напомним, что генетическая дисперсия
существует только тогда, когда имеются
различия в средних значениях признака
у особей с разными генотипами, а в нашем
примере средние совпадают). Таким
образом, несмотря на то, что реальные
генотипы существуют, мы не обнаруживаем
г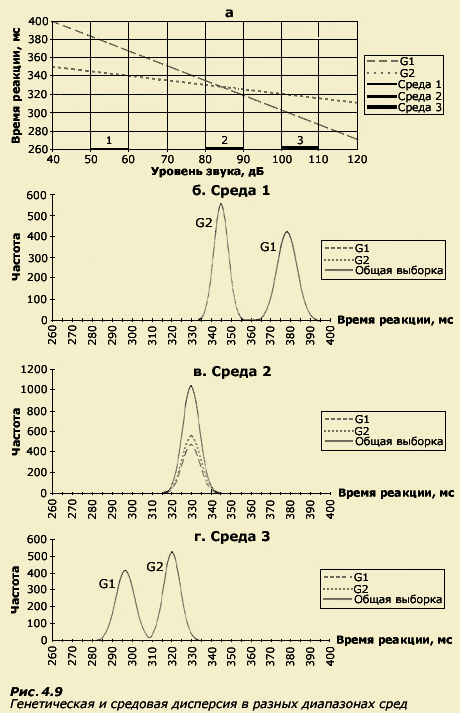 енетической
дисперсии.
Теперь
обратимся к средовым диапазонам 1 и 3
(см. рис. 4.9 б, г). В этих зонах существуют
отчетливые фенотипические различия
между генотипами. В первом диапазоне
среды фенотипические значения признака
для G1 будут всегда выше, чем для G2, тогда
как в третьем диапазоне, наоборот,
значения признака у G1 будут всегда ниже.
В результате общепопуляционное
распределение в средовых диапазонах 1
и 3 будет иметь две отчетливые моды и
значительная часть дисперсии будет
обусловлена генетической составляющей,
поскольку средние по каждому из двух
генотипов заметно отличаются.
На
этом примере мы продемонстрировали
чувствительность показателя наследуемости
к изменению среды. Действительно, в
средовом диапазоне 2 генетическая
дисперсия не обнаруживается, хотя оба
генотипа остаются представленными в
популяции, тогда как при тех же генотипах
в диапазонах 1 и 3 генетическая дисперсия
отчетливо выявляется.
Поскольку
показатель наследуемости определяется
отношением генетической дисперсии к
фенотипической, из этого следует, что
в рассмотренных диапазонах среды
значения показателя наследуемости
будут отличаться. Если в диапазонах 1 и
3 величина h2 будет приближаться
к единице, то в диапазоне 2, наоборот,
будет стремиться к нулю. Отсюда следует
еще одно важное положение количественной
генетики. Мы уже подчеркивали, что
количественное значение показателя
наследуемости не является атрибутом
признака как такового, а относится лишь
к признаку в конкретной популяции с
определенным составом генотипов. К
этому следует добавить, что не только
состав генотипов, но и конкретные
средовые условия, воздействию которых
подвергаются индивиды, определяют
количественные величины генетической
и средовой дисперсии, а следовательно,
и показателя наследуемости - при
помещении популяции в другой диапазон
сред мы можем получить совершенно иной
коэффициент наследуемости.
енетической
дисперсии.
Теперь
обратимся к средовым диапазонам 1 и 3
(см. рис. 4.9 б, г). В этих зонах существуют
отчетливые фенотипические различия
между генотипами. В первом диапазоне
среды фенотипические значения признака
для G1 будут всегда выше, чем для G2, тогда
как в третьем диапазоне, наоборот,
значения признака у G1 будут всегда ниже.
В результате общепопуляционное
распределение в средовых диапазонах 1
и 3 будет иметь две отчетливые моды и
значительная часть дисперсии будет
обусловлена генетической составляющей,
поскольку средние по каждому из двух
генотипов заметно отличаются.
На
этом примере мы продемонстрировали
чувствительность показателя наследуемости
к изменению среды. Действительно, в
средовом диапазоне 2 генетическая
дисперсия не обнаруживается, хотя оба
генотипа остаются представленными в
популяции, тогда как при тех же генотипах
в диапазонах 1 и 3 генетическая дисперсия
отчетливо выявляется.
Поскольку
показатель наследуемости определяется
отношением генетической дисперсии к
фенотипической, из этого следует, что
в рассмотренных диапазонах среды
значения показателя наследуемости
будут отличаться. Если в диапазонах 1 и
3 величина h2 будет приближаться
к единице, то в диапазоне 2, наоборот,
будет стремиться к нулю. Отсюда следует
еще одно важное положение количественной
генетики. Мы уже подчеркивали, что
количественное значение показателя
наследуемости не является атрибутом
признака как такового, а относится лишь
к признаку в конкретной популяции с
определенным составом генотипов. К
этому следует добавить, что не только
состав генотипов, но и конкретные
средовые условия, воздействию которых
подвергаются индивиды, определяют
количественные величины генетической
и средовой дисперсии, а следовательно,
и показателя наследуемости - при
помещении популяции в другой диапазон
сред мы можем получить совершенно иной
коэффициент наследуемости.
4.3.4. Важность правильной интерпретации показателя наследуемости
Поскольку генетическая дисперсия чувствительна к изменениям среды, нужно быть очень осторожными в интерпретации результатов, получаемых в генетике количественных признаков. Если в какой-то популяции мы обнаруживаем высокую генетическую дисперсию признака, т.е. высокую наследуемость, это еще не означает, что менять среду для воздействия на признак не имеет смысла. Действительно, непосвященный человек может рассуждать так: поскольку все различия между индивидами в этой популяции обусловлены различиями в генотипах, а не в условиях среды, мы вряд ли сможем улучшить этот признак, воздействуя на среду. Но такое суждение может быть ошибочным. Достаточно вспомнить только что рассмотренный нами пример, когда переход из одних средовых условий в другие значительно изменяет характеристики популяционной изменчивости и может свести на нет всю генетическую дисперсию. Из сказанного следует: если у нас имеются данные о малой чувствительности признака к среде в каком-то узком средовом диапазоне, это еще не означает, что и в других средовых условиях признак будет нечувствителен к среде. Так же и генотипы, сильно различающиеся по своим проявлениям в одной среде, могут совсем не отличаться в другой. Таким образом, высокая наследуемость ни в коей мере не означает невозможность изменения признака при изменении среды. Этот важный момент особенно часто не понимается, и то, что записано в генах, многие воспринимают как некий рок, фатальную предопределенность. На самом деле это не так: по величине генетической дисперсии и соответственно по величине показателя наследуемости невозможно предсказать последствия изменений среды. Коэффициент наследуемости говорит лишь о том, какая доля генетической изменчивости существует в данной популяции в настоящее время и в существующих условиях среды. Кроме того, необходимо понимать, что показатель наследуемости есть характеристика популяции, а не конкретного индивида и его конкретного фенотипа. Если в каком-то исследовании была получена, например, оценка наследуемости интеллекта, равная 0,6 (60%), мы не должны думать, что у каждого человека интеллект на 60% зависит от генов, а на 40% - от среды. Это довольно распространенное заблуждение. Такой показатель наследуемости лишь означает, что в исследованной популяции в данное время 60% изменчивости по интеллекту возникает за счет разнообразия генотипов и только 40% объясняется разнообразием сред, в которых находятся отдельные индивиды. К фенотипу же отдельного индивида эти показатели не имеют никакого отношения. Методы количественной генетики не позволяют оценить соотношение генетических и средовых факторов в формировании фенотипа конкретного индивида. Как считает один из наиболее известных исследователей в области генетики поведения Роберт Пломин, лучше всего думать, что как наследственность, так и среда одинаково важны для формирования индивидуальности, т.е. можно сказать, что в человеке все на 100% от генов и на 100% от воспитания.
Выводы
Доля генетической составляющей в фенотипической дисперсии признака называется наследуемостью.
Наследуемость не является атрибутом признака как такового, а зависит от состава генотипов в популяции и от конкретных средовых условий.
Высокая наследуемость не означает невозможности изменения признака при изменении среды.
Наследуемость есть характеристика популяции, а не конкретного индивида и его конкретного фенотипа.
4.4. Генотип-средовое взаимодействие
4.4.1. Вклад генотип-средового взаимодействия в популяционную изменчивость
4.4.2. Генотип-средовая ковариация
4.4.1. Вклад генотип-средового взаимодействия в популяционную изменчивость
Очень важно понять,
что два основных источника различий
между людьми (генотип и среда) тесно
связаны друг с другом и находятся в
непрерывном взаимодействии. Различия
между носителями разных генотипов могут
не проявиться, если будут отсутствовать
те факторы среды, которые превращают
генотипические различия в видимые
фенотипические. Мы уже приводили пример
с различиями в цвете кожи в летний и
зимний периоды. Подобных примеров можно
привести множество. Так, люди с генетической
склонностью к полноте вряд ли будут
отличаться тучностью, если в их рационе
не будет достаточного количества
калорийной пищи, например, в условиях
голода или при соблюдении соответствующей
диеты. Люди с исключительными музыкальными
способностями не смогут развить их,
если они не будут иметь возможности для
специальных занятий. То же самое
справедливо для любой области человеческой
деятельности: только в благоприятных
условиях среды люди с особыми природными,
наследственными задатками будут
значительно отличаться от остальной
популяции. Если условия среды мешают
развитию задатков, мы вряд ли получим
большие различия в фенотипах людей,
имеющих разные генотипы. Таким образом,
генотипические различия между людьми
могут не превращаться в фенотипические,
если среда не способствует этому. Это
означает отсутствие фатальной неизбежности
проявления всех генетических задатков,
в том числе и неблагоприятных. Известно,
например, что такое заболевание, как
шизофрения,
имеет значительный наследственный
компонент, однако носители неблагоприятных
генов могут избежать заболевания, если
не будут подвергаться значительным
стрессогенным воздействиям среды,
которые чаще всего и "запускают"
болезнь.
Итак,
имеющиеся между людьми различия в
генотипах могут проявляться или не
проявляться в зависимости от средовых
условий - в одном средовом диапазоне
эти различия будут явно выражены, тогда
как в другом мы и не заподозрим, что они
существуют. Если перевести эти рассуждения
на язык дисперсий, мы можем сказать так:
при одном и том же составе генотипов
могут существовать условия среды,
которые способны менять дисперсию
фенотипов в популяции. Если условия
среды приводят к увеличению фенотипических
различий между носителями разных
генотипов (например, доступность
калорийной пищи в примере со склонностью
к полноте), результатом будет увеличение
фенотипической
дисперсии в популяции.
Если же среда нивелирует различия между
индивидами, фенотипическая дисперсия
будет уменьшаться.
 Итак,
взаимодействие конкретных генотипов
со средой может приводить к изменению
фенотипической дисперсии. Этот феномен
носит название генотип-средового
взаимодействия. Поскольку генотип-средовое
взаимодействие способно влиять на
дисперсию признака в популяции, из этого
следует, что оно порождает определенную
долю фенотипической дисперсии.
Проблема
вклада генотип-средового взаимодействия
в количественную изменчивость начала
обсуждаться достаточно давно. Дж.
Холдейн (Haldane J., 1946; цит. по:
Эрман
Л. и Парсонс П., 1984)
рассматривает такой пример. Допустим,
в популяции существуют два генотипа -
А и В. У этих генотипов в условиях
среды X и Y проводят измерения
какого-то количественного признака.
Полученные количественные значения в
зависимости от степени выраженности
признака обозначают цифрами от 1 до 4.
Число сочетаний факторов А, В, X и Y
равно числу возможных способов
перестановки, т.е. и в данном случае
равняется 24 (4! = 1х2х3х4 = 24). Однако если
условиться, что генотип А в среде X всегда
имеет большие значения признака, чем
генотип В, то остается только шесть
возможных комбинаций, для которых
Дж.Холдейн приводит соответствующую
таблицу. Мы же попробуем изобразить все
шесть ситуаций графически (рис.
4.10).
Рассмотрим
первый и второй графики. Мы видим, что
и в первой, и во второй средах генотип
А имеет более высокие значения признака,
чем генотип В, причем прямые параллельны,
а это означает, что дисперсия, т.е. разброс
значений фенотипов, во всех средах
остается одинаковой. Таким образом,
генотип-средовое взаимодействие как
компонента дисперсии отсутствует.
Теперь посмотрим, имеется ли реальное
взаимодействие генотипа и среды в
соответствии с представлениями о норме
реакции. Мы видим, что при переходе из
среды X в среду Y для каждого из
генотипов произошло изменение
количественных значений признака. При
этом генотипы отреагировали на изменение
среды сходным образом, сохранив свои
относительные положения. На этом примере
ясно видно, что взаимодействие генотипа
и среды может пониматься двояко. С одной
стороны, существует реальное взаимодействие
каждого генотипа со средой, приводящее
к изменению конкретного фенотипа. С
другой стороны, имеется эффект
статистического взаимодействия факторов
наследственности и среды, влияющий на
величину соответствующего компонента
дисперсии. В данном случае, несмотря на
существование реального взаимодействия
конкретных генотипов и конкретных
условий среды, компонента фенотипической
дисперсии, обусловленная взаимодействием
двух факторов, оказалась равной нулю.
Этот момент необходимо четко уяснить,
поскольку очень часто реальное
взаимодействие генотипа и среды
смешивается со статистическим эффектом
взаимодействия факторов.
Первый
и второй графики сходны, различаются
лишь направления изменения фенотипов
при переходе из среды X в среду Y.
Для первого варианта примером может
служить такая реальная ситуация: ученики
младших классов, обучавшиеся у молодого
неопытного учителя, попадают к сильному
педагогу, и при этом все улучшают свои
показатели, но сохраняют те же ранговые
места. Обратная ситуация может быть
примером для второго графика. Можно
представить себе и такую ситуацию. Люди
с высокой и низкой тревожностью будут
сходно реагировать на ситуацию
неопределенности и тревоги ухудшением
показателей деятельности, но деятельность
первых будет страдать сильнее, чем
деятельность вторых.
Третий
график отличается от первых двух тем,
что линии здесь пересекаются. Соответственно
по своим фенотипическим значениям в
среде X генотип А будет опережать В, а в
среде Y - наоборот. Оба генотипа при
переходе из среды X в среду Y будут давать
прирост фенотипических значений
признака, но в разной мере. Генотип В
оказывается более чувствительным к
среде, соответственно он дает и больший
прирост, сразу на четыре единицы, тогда
как генотип А - всего на две. В этом случае
генотип-средовое взаимодействие как
компонента дисперсии будет отличным
от нуля. Примером такого рода взаимодействия
может послужить такая ситуация, когда
общительный (А) и необщительный (В)
ребенок во время болезни находятся дома
(среда У) или в больнице (среда X). Дети
более комфортно чувствуют себя в домашних
условиях, в то же время общительного
ребенка вынужденная изоляция от
сверстников тяготит, однако в больнице
он будет лучше адаптироваться, чем
необщительный ребенок.
На
четвертом графике генотип А в обоих
средах имеет большие фенотипические
значения признака, но генотипы по-разному
реагируют на переход из среды X в среду
Y. Для генотипа А характерно уменьшение
значений, а для генотипа В - увеличение.
Здесь также присутствует генотип-средовое
взаимодействие как компонент дисперсии.
Воспользуемся примером, который приводит
Дж.
Холдейн. Он рассматривает
нормальных (А) и умственно отсталых (В)
детей, которые обучаются в обычной
(среда X) и специализированной (среда Y)
школах. Понятно, что в обычной школе
умственно отсталые дети будут значительно
отставать от обычных детей, тогда как
в специализированной школе с программой,
рассчитанной на умственно отсталых
детей, последние дадут прирост успешности,
а нормальные дети будут отставать от
своих сверстников.
На
пятом графике опять наблюдается
пересечение линий. При переходе из среды
X в среду Y генотип А демонстрирует
уменьшение значений, тогда как генотип
В - увеличение. В среде X фенотипические
различия между генотипами больше, чем
в среде Y. Опять мы видим, что генотип-средовое
взаимодействие меняет дисперсию
фенотипов. В качестве примера можно
рассмотреть людей с высоким (А) и низким
(В) уровнями активации. В условиях тишины
и изоляции (X) более эффективно будут
работать первые, тогда как в условиях
шума (Y) - вторые.
И
последний, шестой вариант. Линии на
графике опять пересекаются. Генотип В
более чувствителен к изменению среды
и демонстрирует рост величины признака
при переходе в среду Y, тогда как генотип
А менее чувствителен к среде и реагирует
иначе. Генотип-средовое взаимодействие
вносит вклад в дисперсию признака.
Возможным примером может служить
ситуация, в которой генотипам А и В
соответствуют дети с разными типами
способностей. Одни дети (генотип А)
обладают средним уровнем способностей,
при опережающем развитии пространственных
способностей, а другие (генотип В) имеют
ярко выраженные вербальные способности
и испытывают серьезные затруднения при
решении пространственных задач. В
соответствии с этим дети будут
демонстрировать разную успешность на
уроках геометрии (X) и литературы (Y). Все
эти примеры наглядно демонстрируют,
что изменчивость в популяции складывается
не только за счет простого суммирования
эффектов среды и генотипа, но и включает
компонент генотип-средового взаимодействия.
Наличие генотип-средового взаимодействия
увеличивает популяционную дисперсию
признака. Это связано с тем, что
представители различных генотипов
по-разному реагируют на одни и те же
изменения среды. Это и приводит к
увеличению популяционной изменчивости.
Чтобы
обнаружить статистический компонент
Г-С взаимодействия необходимо подобрать
группы индивидов, отличающихся своими
генетическими конституциями, и находящихся
в различных типах условий среды. В
психогенетических исследованиях, как
правило, так и поступают. Например, при
исследованиях аффективных расстройств
(Kendler K.S et al., 1995, цит. по Равич-Щербо И.В.
и др., 1999) можно подобрать группы испытуемых
из генетически отягощенных и неотягощенных
семей. Это и будут контрастные генотипы
("плохой" и "хороший"). Внутри
каждой из этих групп можно подобрать
индивидов, находящихся в контрастных
средах (благоприятной и неблагоприятной).
Таким образом, мы будем иметь 4 группы:
в первой из них "хороший" генотип
будет сочетаться с "хорошей" средой,
во второй - "хороший" генотип будет
сочетаться с "плохой" средой, в
третьей - "плохой" генотип будет
находиться в "хорошей" среде и,
наконец, в четвертой - и генотип, и среда
будут "плохими" (табл. 4.3).
Итак,
взаимодействие конкретных генотипов
со средой может приводить к изменению
фенотипической дисперсии. Этот феномен
носит название генотип-средового
взаимодействия. Поскольку генотип-средовое
взаимодействие способно влиять на
дисперсию признака в популяции, из этого
следует, что оно порождает определенную
долю фенотипической дисперсии.
Проблема
вклада генотип-средового взаимодействия
в количественную изменчивость начала
обсуждаться достаточно давно. Дж.
Холдейн (Haldane J., 1946; цит. по:
Эрман
Л. и Парсонс П., 1984)
рассматривает такой пример. Допустим,
в популяции существуют два генотипа -
А и В. У этих генотипов в условиях
среды X и Y проводят измерения
какого-то количественного признака.
Полученные количественные значения в
зависимости от степени выраженности
признака обозначают цифрами от 1 до 4.
Число сочетаний факторов А, В, X и Y
равно числу возможных способов
перестановки, т.е. и в данном случае
равняется 24 (4! = 1х2х3х4 = 24). Однако если
условиться, что генотип А в среде X всегда
имеет большие значения признака, чем
генотип В, то остается только шесть
возможных комбинаций, для которых
Дж.Холдейн приводит соответствующую
таблицу. Мы же попробуем изобразить все
шесть ситуаций графически (рис.
4.10).
Рассмотрим
первый и второй графики. Мы видим, что
и в первой, и во второй средах генотип
А имеет более высокие значения признака,
чем генотип В, причем прямые параллельны,
а это означает, что дисперсия, т.е. разброс
значений фенотипов, во всех средах
остается одинаковой. Таким образом,
генотип-средовое взаимодействие как
компонента дисперсии отсутствует.
Теперь посмотрим, имеется ли реальное
взаимодействие генотипа и среды в
соответствии с представлениями о норме
реакции. Мы видим, что при переходе из
среды X в среду Y для каждого из
генотипов произошло изменение
количественных значений признака. При
этом генотипы отреагировали на изменение
среды сходным образом, сохранив свои
относительные положения. На этом примере
ясно видно, что взаимодействие генотипа
и среды может пониматься двояко. С одной
стороны, существует реальное взаимодействие
каждого генотипа со средой, приводящее
к изменению конкретного фенотипа. С
другой стороны, имеется эффект
статистического взаимодействия факторов
наследственности и среды, влияющий на
величину соответствующего компонента
дисперсии. В данном случае, несмотря на
существование реального взаимодействия
конкретных генотипов и конкретных
условий среды, компонента фенотипической
дисперсии, обусловленная взаимодействием
двух факторов, оказалась равной нулю.
Этот момент необходимо четко уяснить,
поскольку очень часто реальное
взаимодействие генотипа и среды
смешивается со статистическим эффектом
взаимодействия факторов.
Первый
и второй графики сходны, различаются
лишь направления изменения фенотипов
при переходе из среды X в среду Y.
Для первого варианта примером может
служить такая реальная ситуация: ученики
младших классов, обучавшиеся у молодого
неопытного учителя, попадают к сильному
педагогу, и при этом все улучшают свои
показатели, но сохраняют те же ранговые
места. Обратная ситуация может быть
примером для второго графика. Можно
представить себе и такую ситуацию. Люди
с высокой и низкой тревожностью будут
сходно реагировать на ситуацию
неопределенности и тревоги ухудшением
показателей деятельности, но деятельность
первых будет страдать сильнее, чем
деятельность вторых.
Третий
график отличается от первых двух тем,
что линии здесь пересекаются. Соответственно
по своим фенотипическим значениям в
среде X генотип А будет опережать В, а в
среде Y - наоборот. Оба генотипа при
переходе из среды X в среду Y будут давать
прирост фенотипических значений
признака, но в разной мере. Генотип В
оказывается более чувствительным к
среде, соответственно он дает и больший
прирост, сразу на четыре единицы, тогда
как генотип А - всего на две. В этом случае
генотип-средовое взаимодействие как
компонента дисперсии будет отличным
от нуля. Примером такого рода взаимодействия
может послужить такая ситуация, когда
общительный (А) и необщительный (В)
ребенок во время болезни находятся дома
(среда У) или в больнице (среда X). Дети
более комфортно чувствуют себя в домашних
условиях, в то же время общительного
ребенка вынужденная изоляция от
сверстников тяготит, однако в больнице
он будет лучше адаптироваться, чем
необщительный ребенок.
На
четвертом графике генотип А в обоих
средах имеет большие фенотипические
значения признака, но генотипы по-разному
реагируют на переход из среды X в среду
Y. Для генотипа А характерно уменьшение
значений, а для генотипа В - увеличение.
Здесь также присутствует генотип-средовое
взаимодействие как компонент дисперсии.
Воспользуемся примером, который приводит
Дж.
Холдейн. Он рассматривает
нормальных (А) и умственно отсталых (В)
детей, которые обучаются в обычной
(среда X) и специализированной (среда Y)
школах. Понятно, что в обычной школе
умственно отсталые дети будут значительно
отставать от обычных детей, тогда как
в специализированной школе с программой,
рассчитанной на умственно отсталых
детей, последние дадут прирост успешности,
а нормальные дети будут отставать от
своих сверстников.
На
пятом графике опять наблюдается
пересечение линий. При переходе из среды
X в среду Y генотип А демонстрирует
уменьшение значений, тогда как генотип
В - увеличение. В среде X фенотипические
различия между генотипами больше, чем
в среде Y. Опять мы видим, что генотип-средовое
взаимодействие меняет дисперсию
фенотипов. В качестве примера можно
рассмотреть людей с высоким (А) и низким
(В) уровнями активации. В условиях тишины
и изоляции (X) более эффективно будут
работать первые, тогда как в условиях
шума (Y) - вторые.
И
последний, шестой вариант. Линии на
графике опять пересекаются. Генотип В
более чувствителен к изменению среды
и демонстрирует рост величины признака
при переходе в среду Y, тогда как генотип
А менее чувствителен к среде и реагирует
иначе. Генотип-средовое взаимодействие
вносит вклад в дисперсию признака.
Возможным примером может служить
ситуация, в которой генотипам А и В
соответствуют дети с разными типами
способностей. Одни дети (генотип А)
обладают средним уровнем способностей,
при опережающем развитии пространственных
способностей, а другие (генотип В) имеют
ярко выраженные вербальные способности
и испытывают серьезные затруднения при
решении пространственных задач. В
соответствии с этим дети будут
демонстрировать разную успешность на
уроках геометрии (X) и литературы (Y). Все
эти примеры наглядно демонстрируют,
что изменчивость в популяции складывается
не только за счет простого суммирования
эффектов среды и генотипа, но и включает
компонент генотип-средового взаимодействия.
Наличие генотип-средового взаимодействия
увеличивает популяционную дисперсию
признака. Это связано с тем, что
представители различных генотипов
по-разному реагируют на одни и те же
изменения среды. Это и приводит к
увеличению популяционной изменчивости.
Чтобы
обнаружить статистический компонент
Г-С взаимодействия необходимо подобрать
группы индивидов, отличающихся своими
генетическими конституциями, и находящихся
в различных типах условий среды. В
психогенетических исследованиях, как
правило, так и поступают. Например, при
исследованиях аффективных расстройств
(Kendler K.S et al., 1995, цит. по Равич-Щербо И.В.
и др., 1999) можно подобрать группы испытуемых
из генетически отягощенных и неотягощенных
семей. Это и будут контрастные генотипы
("плохой" и "хороший"). Внутри
каждой из этих групп можно подобрать
индивидов, находящихся в контрастных
средах (благоприятной и неблагоприятной).
Таким образом, мы будем иметь 4 группы:
в первой из них "хороший" генотип
будет сочетаться с "хорошей" средой,
во второй - "хороший" генотип будет
сочетаться с "плохой" средой, в
третьей - "плохой" генотип будет
находиться в "хорошей" среде и,
наконец, в четвертой - и генотип, и среда
будут "плохими" (табл. 4.3).
Табл. 4.3.
Схема эксперимента
|
Неотягощенный генотип (G+) |
Отягощенный генотип (G-) |
Благоприятная среда (Е+) |
G+ E+ |
G- E+ |
Неблагоприят-ная среда (Е-) |
G+ E- |
G- E- |
Это классический пример планирования эксперимента для модели двухфакторного дисперсионного анализа с целью выявления эффекта взаимодействия факторов. Если в таком эксперименте мы выявим разную чувствительность контрастных генотипов к средовым изменениям, то можно делать заключение о наличии Г-С взаимодействия (рис. 4.11).

Г-С взаимодействие легко обнаруживается в экспериментах на животных. Классическим примером является разная обучаемость в лабиринте селектированных "умных" и "глупых" крыс в зависимости от условий выращивания молодняка (рис. 4.12)
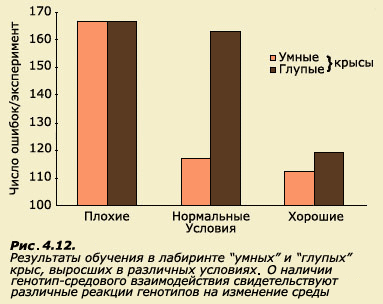
Механизм увеличения фенотипической дисперсии при наличии Г-С взаимодействия можно наглядно продемонстрировать на примере депрессивного расстройства, которое можно отнести к разряду психических заболеваний с наследственной предрасположенностью и пороговым эффектом. В популяции существуют индивиды с различной наследственной предрасположенностью к заболеванию (рис 4. 13).
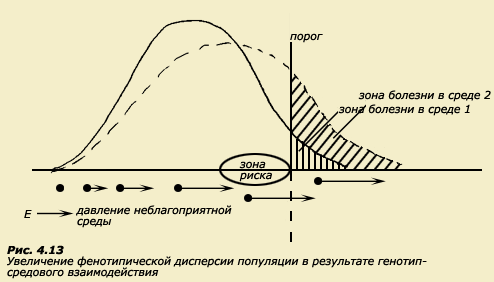
Часть из них находится в острой стадии болезни, т.е. за условным порогом (правая часть распределения за чертой). Предрасположенные из группы риска имеют гораздо больше шансов оказаться за порогом болезни, т.е. попасть в группу больных. Вместе с тем имеются индивиды, абсолютно не подверженные депрессии - они располагаются в противоположном конце распределения (слева). Допустим, вся популяция начинает подвергаться неблагоприятным условиям среды (E - обозначено стрелкой внизу слева), способствующим развитию депрессии (например, увеличивается стрессогенность среды). Тогда можно предположить, что индивиды, устойчивые к развитию депрессии, совсем не изменят свой фенотип, т.е. не будут проявлять даже малейших признаков депрессии. Это означает, что левая часть распределения не будет смещаться в сторону болезни. Что же будет происходить с остальными представителями популяции? Можно ожидать, что чувствительность к стрессогенной среде будет нарастать по мере приближения к порогу, и индивиды будут проявлять тем более признаков депрессии в своем фенотипе, чем ближе они к группе риска. Смещение фенотипов на графике показано стрелками разной длины. Чем более чувствителен генотип, тем длиннее стрелка. Таким образом, из рисунка становится понятно, что все распределение при переходе популяции в неблагоприятную среду начинает как бы растягиваться, причем левый его край стоит на месте, а правый - сдвигается в сторону болезни. Пунктирная линия на графике представляет собой новое распределение фенотипов в измененной среде: оно более растянуто, что говорит об увеличении фенотипической дисперсии; кроме того, заметно, что гораздо больше индивидов окажется в зоне болезни (за пороговой чертой). Этот пример демонстрирует, что при разной реакции генотипов на изменение среды фенотипическая дисперсия меняется именно за счет компонента Г-С взаимодействия. В предыдущих темах мы рассматривали упрощенные модели, в которых принималось, что влияния генотипа и среды аддитивны. В таких случаях фенотипическая дисперсия представляет собой сумму генетической и средовой компоненты (VP = VG + VE). Если же имеют место Г-С взаимодействия, то в формулу фенотипической структуры популяции должен быть добавлен еще один компонент (VGE - дисперсия, обусловленная генотип-средовым взаимодействием). Тогда она примет следующий вид:
VP = VG + VE + VGE.
О наличии Г-С взаимодействия свидетельствуют и различия в значениях коэффициентов наследуемости в контрастных средах. Например, при изучении наследуемости алкоголизма (Heath A.S. et al., 1989, цит. по Равич-Щербо И.В. и др., 1999) было обнаружено, что в популяции замужних женщин он оказывается ниже, чем в популяции незамужних. Более высокий коэффициент наследуемости говорит о более ярком проявлении генетической вариативности в среде, соответствующей ситуации незамужней женщины. Следовательно, в ситуации замужества наследственные различия по предрасположенности к алкоголизму, по-видимому, не проявляются столь заметно в фенотипических различиях. Из этого можно заключить, что отягощенный генотип должен быть более чувствителен к изменениям среды (рис. 4.14).
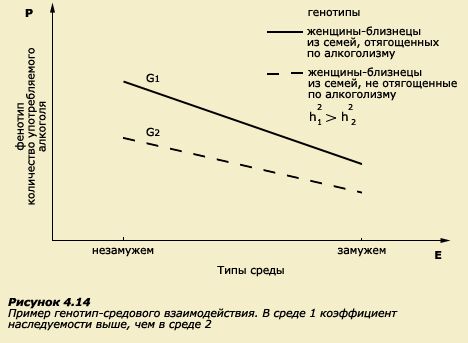
Еще раз подчеркнем, что следует различать генотип-средовое взаимодействие в статистическом смысле и реальное взаимодействие генотипа и среды при формировании конкретного фенотипа.
4.4.2. Генотип-средовая ковариация
Рассмотренное нами генотип-средовое взаимодействие следует отличать от другого явления, с которым его часто путают. Речь идет о генотип-средовой ковариации (или корреляции). Генотип-средовая ковариация имеет место тогда, когда генотипы оказываются в определенных средах не случайно, а в соответствии со своей наследственной предрасположенностью. Это означает, что генотипы неравномерно распределяются по разным средам. Примером может служить соответствие различных видов, рас, подвидов, разновидностей различным местам обитания. Приспособленность генотипов к определенным условиям среды приводит к тому, что генотипы концентрируются в тех средах, которые больше соответствуют их особенностям. В человеческих популяциях наблюдаются похожие явления. В таких различных средах, как стадион, концертный зал или библиотека, мы, скорее всего, встретим людей с определенными генетическими предрасположенностями: на стадионе будут чаще встречаться люди с природной склонностью к двигательной деятельности, в концертном зале - люди с музыкальной одаренностью, а в библиотеке будет больше людей с высокими умственными способностями. Мы видели, что генотип-средовое взаимодействие всегда увеличивает популяционную дисперсию. Генотип-средовая ковариация может как увеличивать, так и уменьшать дисперсию фенотипов в популяции. В жизни человека генотип-средовые ковариации играют важную роль. Родители могут передавать ребенку не только определенную генетическую предрасположенность, но и одновременно обеспечивать ему соответствующую среду для развития природных задатков. Здесь мы имеем дело с культурной преемственностью, которую не всегда легко отличить от наследственности. Например, родители-музыканты большей частью заботятся о том, чтобы их дети также получили музыкальное образование. В таких семьях можно ожидать, что дети вместе с "генами музыкальности" унаследуют от родителей и "музыкальную среду". Генотип-средовая ковариация, как и генотип-средовое взаимодействие, влияет на общую дисперсию признака, а следовательно, может влиять на величину оценок наследуемости. Это следует учитывать при проведении исследований в генетике поведения. Рассмотрим явление генотип-средовой ковариации более подробно. Сначала обратимся к математике, чтобы понять, что генотип-средовая ковариация является компонентом дисперсии фенотипов. Мы знаем, что фенотип представляет собой результат совместного действия генотипа и среды. Если перевести это на язык статистики, то количественное значение признака у конкретного i-того индивида можно представить как фенотипическое отклонение (Pi) от популяционной средней. Такое значение фенотипа есть результат, с одной стороны, генотипического отклонения (Gi) от популяционной средней и, с другой стороны, - средового отклонения (Ei). Таким образом, можно записать языком математики, что конкретный фенотип есть результат взаимодействия генотипа и среды:
Pi = Gi + Ei.
Чтобы определить дисперсии отклонений, нужно каждое i-е отклонение возвести в квадрат, просуммировать все квадраты отклонений и суммы разделить на n.
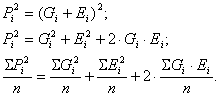
Величина
![]() представляет
собой общую фенотипическую дисперсию,
представляет
собой общую фенотипическую дисперсию,
![]() -
генетическая дисперсия,
-
генетическая дисперсия,
![]() -
средовая дисперсия. Поскольку Gi
и Ei представляют собой генотипические
и средовые отклонения, то сумма их
произведений
-
средовая дисперсия. Поскольку Gi
и Ei представляют собой генотипические
и средовые отклонения, то сумма их
произведений
![]() есть
ковариация генотипа и среды. Итак,
есть
ковариация генотипа и среды. Итак,
VP = VG + VE + 2covGE.
Если считать, что генотипы равномерно распределены по средам (т.е. оказываются в определенной среде совершенно случайно), то компонент 2covGE становится равным 0, и тогда мы опять имеем ту формулу фенотипической структуры популяции, которая рассматривалась нами как исходная:
VP = VG + VE.
В реальных популяциях генотип-средовая ковариация является обычным явлением и ее вклад в вариативность признаков, как правило, не равен нулю. Действительно, трудно себе представить, чтобы индивид совершенно случайно попадал в ту или иную среду. С рождением каждого ребенка популяция пополняется еще одним генотипом, но ребенок обычно сразу же попадает в среду своей собственной семьи, к своим собственным родителям, бабушкам, дедушкам, братьям и сестрам, с которыми у него есть общие гены. Если при игре в рулетку брошенный шарик случайно оказывается в ячейке, то только что родившийся ребенок занимает "ячейку" вполне определенную: в своей семье, своем городе, своей стране. Таким образом, его среда не может быть совершенно случайной. Рассмотрим следующую схему (рис. 4.15).
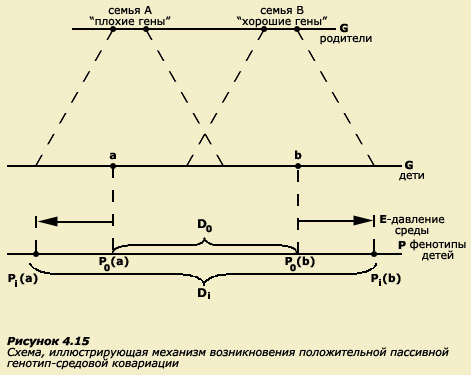
Предположим, в популяции существуют две семьи (А и В). Супруги А, оба, обладают "плохими" генами в отношении какого-то количественного признака с аддитивным типом наследования (допустим, это интеллект), т.е. у них преобладают гены-ослабители. В то же время в другой семье (В) супруги имеют "хорошие" гены (преобладают гены-усилители). В результате рекомбинаций при образовании гамет дети супругов А и супругов В могут получить различные сочетания генов, и, в принципе, среди них могут оказаться обладатели сходных сочетаний (зона перекрытия на рисунке), однако все же вероятность того, что дети супругов А окажутся обладателями "худших" генов достаточно велика, и, наоборот, дети супругов В скорее всего получат "лучшие" гены. На схеме генотипы детей обозначены точками а и b на соответствующей оси (внизу). Теперь представим себе, в какой среде окажутся дети в этих семьях. Скорее всего в силу более высоких умственных способностей родители В обеспечат своему ребенку лучшую среду для развития, чем родители А. В семье В среда будет обогащенной (больше книг, развивающих игрушек, дополнительных занятий, речевое общение более высокого уровня и т.д.), а в семье А - обедненной, поскольку у родителей, вероятно, будет меньше возможностей создать подобную среду. Следовательно, среда семьи В будет способствовать максимальному развитию исходных задатков своего ребенка, и его фенотип будет постоянно улучшаться. В то же время среда семьи А не создаст возможностей для развития фенотипа ребенка. В результате наследственно заложенные задатки не проявятся в полной степени. Таким образом, мы имеем ситуацию, в которой совершенно пассивно, просто по факту своего рождения, обладатель лучшего генотипа унаследовал и лучшую среду, а обладатель худшего генотипа унаследовал худшую среду. Здесь мы имеем дело с явлением, которое в психогенетике называется пассивной положительной ковариацией генотипа и среды. Ковариация (корреляция) всегда положительна, если два параметра изменяются (ковариируют) в одном направлении. Например, чем больше рост, тем больше длина ступни человека. Это пример положительной ковариации. В рассмотренном нами случае генотип и среда также ковариируют положительно, поскольку лучшему генотипу соответствует лучшая среда и наоборот. Пассивной ковариация генотипа и среды называется потому, что ни родители, ни дети не прикладывают специальных усилий для создания среды: дети просто "наследуют" среду так же, как и гены. Это называется культурным наследованием. На той же схеме (рис. 4.15) стрелками показано, что среда как бы "тянет" фенотипы (Pa) и (Pb) в разные стороны. Понятно, что в результате та разница, которая имелась между детьми при рождении (диапазон фенотипических различий - D0), будет усугубляться (диапазон фенотипических различий - Di). Если такая картина наблюдается не только в одной семье, а типична для популяции вообще, то результатом генотип-средовой ковариации будет увеличение фенотипической дисперсии, поскольку разброс между фенотипами в результате такой ковариации, как мы видим, возрастает. Теперь рассмотрим еще одну подобную схему (рис. 4.16).
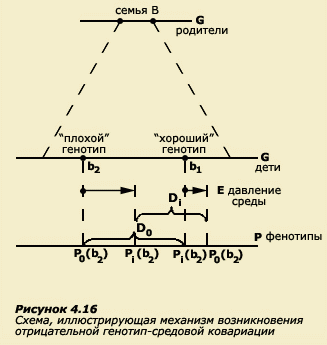
Предположим, что в семье В родился второй ребенок (b2), которому "не повезло", и он получил худшее сочетание генов. Родители обнаружили, что второй ребенок не столь одарен от природы, как первый, и направили все усилия на его развитие, забросив первого, который "и сам пробьется". Тем самым они создали худшему генотипу лучшие условия среды. Это пример отрицательной генотип-средовой ковариации, поскольку переменные варьируют в различных направлениях (хуже генотип - лучше среда и, наоборот, лучше генотип - хуже среда). В результате фенотипические различия между детьми начнут уменьшаться (конечный диапазон различий Di меньше начального - D0). Таким образом, отрицательная генотип-средовая ковариация должна уменьшать популяционную дисперсию, но это в том случае, если рассмотренный пример представляет собой типичное явление. Напоминаем, что ковариация является величиной статистической и может быть обнаружена только на значительных выборках. Если же подобные усилия родителей наблюдаются лишь в одной семье, а в других происходит нечто иное, то никакой отрицательной ковариации генотипа и среды мы не получим. Только что рассмотренная ситуация является примером отрицательной реактивной ковариации генотипа и среды. Реактивной обычно называется ковариация, возникающая в результате специальных усилий социального окружения ребенка в ответ на какие-либо его специфические особенности, связанные с наследственной конституцией. Например, дети, проявляющие признаки природной одаренности в спорте, музыке, изобразительной деятельности, привлекают внимание взрослых (родителей, педагогов, воспитателей), и те стараются создать условия для дальнейшего развития природных задатков: помещают детей в специализированные школы, кружки по интересам и т.п. В результате "хороший" генотип получает "хорошую" среду, т.е. наблюдается положительная ковариация. Реактивная ковариация может быть и отрицательной. Она может возникать, например, в том случае, когда наследственной отягощенности или какому-либо неблагополучию ребенка, обусловленному наследственными причинами, противопоставляются компенсирующие усилия родителей, врачей, педагогов, заставляющие работать среду против генотипа. Однако это скорее исключение, чем правило. Большинство детей с проблемами развития оказываются в среде, которая только усугубляет их неблагополучие. Например, большие трудности часто возникают у детей с гиперактивностью и нарушениями внимания, нарушениями эмоционально-волевой сферы, "трудным" темпераментом. Все это можно назвать синдромом "трудного" ребенка. К сожалению, такие "особые" дети чаще вызывают отрицательные реакции окружающих: их больше наказывают, унижают и обижают, они реже бывают успешными в различных видах деятельности. Получается, что их природное (чаще всего оно и наследственное) неблагополучие влечет за собой и неблагоприятную среду. Ковариация генотипа и среды в этом случае будет положительной, поскольку и генотип, и среда действуют в одном направлении ("плохой" генотип влечет за собой "плохую" среду). Безусловно, это приводит к отрицательным последствиям для фенотипа, но генотип-средовая ковариация в данном случае положительная. Усилия практических психологов должны быть направлены на преодоление существующей тенденции. Необходимо помогать родителям, воспитателям и педагогам создавать такие условия среды, которые будут улучшать фенотип ребенка, имеющего ту или иную отягощенность. Практический психолог, участвуя в отборе детей в специализированные школы, консультируя родителей, участвуя в организации среды в детских учреждениях, должен помнить, что от его усилий зависит, произойдут ли положительные сдвиги в развитии ребенка. Поэтому при "плохом" генотипе следует создавать отрицательную ковариацию генотипа и среды, а при "хорошем" - положительную. Мы рассмотрели два типа ковариации - пассивную и реактивную - и обнаружили, что она может быть как положительной, так и отрицательной. Если вернуться к формуле:
VP = VG + VE +2covGE,
то становится понятным, что положительная ковариация должна увеличивать фенотипическую дисперсию в популяции, а отрицательная - уменьшать. В психогенетике, помимо пассивной и реактивной форм ковариации генотипа и среды, выделяют еще и активную ковариацию. Она характерна для более старших возрастов, начиная с подросткового, когда индивид самостоятельно (активно) выбирает ту среду, которая ему более подходит. Часто это происходит против воли родителей и других взрослых. Типичным примером активной генотип-средовой ковариации является судьба самого Ф. Гальтона, который по настоянию отца долгое время готовился к карьере врача, но после смерти отца забросил медицину и хотя и продолжал изучать природу человека, но уже в совершенно ином аспекте. Таким образом, получается, что не столько среда влияет на формирование индивидуальности человека, сколько сама индивидуальность (с ее уникальной генетической конституцией) формирует свою особую среду. Впервые на это обстоятельство обратила внимание американская исследовательница Сандра Скарр в 1983 г. (Scarr S., McCartney K., 1983). Наличие генотип-средовой ковариации приводит к тому, что средовые условия у более близких родственников, имеющих высокую долю общих генов (т.е. сходные генотипы), оказываются более сходными, чем у более отдаленных родственников. Характерным примером могут служить идентичные близнецы, разлученные в раннем детстве и никогда не встречавшиеся друг с другом. Уже будучи солидными людьми, при первой встрече они с удивлением обнаруживают множество совпадающих обстоятельств своей жизни: сходство профессий, привычек, любимых занятий и даже сходные особенности спутников жизни. Можно ли оценить вклад генотип-средовой ковариации в изменчивость признака? В психогенетике для этого существуют специальные методы. Большинство из них используют приемы, основанные на сравнении оценок характеристик среды у родственников различной степени родства. Одним из наиболее распространенных является методика интервью и наблюдений НОМЕ (Home Observation of Measurement of the Environment), позволяющая оценить эмоциональные и вербальные реакции родителей, отношение к наказаниям, организацию физической среды в семье и т.п. Используются также всевозможные опросники, в которых родственники оценивают различные параметры семейной и внесемейной среды. Полученные оценки могут сравниваться у родственников подобно тому, как это делается для любых количественных характеристик фенотипа. Так можно оценить сходство родственников не только по фенотипу, но и по средовым параметрам. Если средовое сходство оказывается выше у более близких родственников, можно предполагать наличие генотип-средовой ковариации. (В качестве дополнительного материала см. хрестоматии 4.1. и 4.2.)
Выводы
4.1. Количественная изменчивость и методы ее описания
Требования к психологическим измерениям в генетике поведения соответствуют основным требованиям психометрики (надежность, валидность, репрезентативность).
Распределение частот встречаемости различных количественных значений признака в популяции характеризуются двумя статистическими величинами - центральной тенденцией (мода, медиана, среднее) и разбросом значений вокруг среднего (дисперсия).
Дисперсия характеризует межиндивидуальные различия (изменчивость, вариативность).
Генетика поведения изучает природу индивидуальных различий.
4.2. Наследственность и среда как факторы возникновения количественной изменчивости
Вся совокупность генов организма составляет его генотип.
Любые проявления организма в каждый момент жизни составляют его фенотип.
Фенотип есть результат взаимодействия генотипа со средой.
Количественная изменчивость может возникать в результате полимерного действия многих генов на один признак.
Дисперсия количественного признака, возникающая за счет действия генов, носит название генетической дисперсии.
Существуют различные формы взаимодействия генов: аддитивное, доминирование, эпистаз и др.
Количественная изменчивость может возникать под действием факторов среды.
Дисперсия количественного признака, возникающая за счет средовых влияний, носит название средовой дисперсии.
Генотипы по-разному реагируют на одни и те же изменения среды. Специфический характер реакции данного гeнотипа на изменение окружающих условий носит название нормы реакции.
Генотипы отличаются по своей чувствительности к среде.
Фенотипическая изменчивость в популяции складывается из генетической и средовой изменчивости; фенотипическая популяционная дисперсия признака представляет собой сумму его генетической и средовой дисперсий.
4.3. Показатель наследуемости и его особенности
Доля генетической составляющей в фенотипической дисперсии признака называется наследуемостью.
Наследуемость не является атрибутом признака как такового, а зависит от состава генотипов в популяции и от конкретных средовых условий.
Высокая наследуемость не означает невозможности изменения признака при изменении среды.
Наследуемость есть характеристика популяции, а не конкретного индивида и его конкретного фенотипа.