

Группа крови АВ:
гетерозиготы - IА IB.
Люди с группой крови 0 - это всегда гомозиготы, люди с группой крови АВ - это всегда гетерозиготы, а среди людей с группами А и В могут встречаться и гомо- и гетерозиготы. Доминирования в данном случае не наблюдается, но нет и промежуточных форм, и люди с типом крови АВ одновременно проявляют признаки группы А и группы В (Хрестомат. 3.4).
3.4.2. Генные мутации
Несмотря на большую точность репликации ДНК во время процессов митоза и мейоза, время от времени неизбежно возникают ошибки, которые приводят к изменению последовательности нуклеотидов в цепи ДНК, или генным мутациям. Мутация может представлять собой замену одной пары оснований на другую, потерю одного или нескольких нуклеотидов или, наоборот, добавление дополнительных нуклеотидов. При этом худшим вариантом будет потеря или добавление одного или двух нуклеотидов. В этих случаях рамка считывания триплетов неизбежно сдвигается на одно-два основания вправо или влево и все последующие триплеты будут считываться неправильно. Если же выпали или добавились сразу три основания, то изменения коснутся лишь одной аминокислоты, а вся остальная цепь останется верной. Генные мутации, возникающие в гаметах, оказывают разнообразное действие на организм. Многие из них летальны, так как вызывают слишком серьезные нарушения развития. Известно, например, что у человека около 20% беременностей заканчиваются естественным выкидышем в сроки до 12 недель, и в половине этих случаев обнаруживаются аномалии наследственного аппарата, обусловленные, впрочем, не только генными мутациями. Генная мутация может привести к тому, что данному локусу будет соответствовать несколько аллелей. Это увеличивает генетическое разнообразие и повышает количество гетерозиготных особей. Предполагается, что все генетические полиморфизмы возникли из-за замены, потери или добавления нуклеотидов в процессе эволюции. Большинство генных мутаций рецессивны по отношению к "нормальному" аллелю. Такие мутантные аллели могут циркулировать в популяции в течение многих поколений, пока им не удастся встретиться и проявиться. Время от времени могут возникать и доминантные мутантные аллели, которые сразу же дают эффект. Мутации, происходящие в соматических клетках, наследуются только теми клетками, которые образуются из мутантной клетки путем митоза. Они могут повлиять только на тот организм, в котором они возникли, но со смертью особи исчезают из генофонда популяции. При некоторых соматических мутациях возникают клетки с повышенной скоростью роста, что приводит к возникновению опухолей.
3.4.3. Хромосомные аномалии
Помимо генных перестроек, генных мутаций, возможны перестройки и на уровне хромосом, которые отличаются от нормальной рекомбинации, происходящей в мейозе. Хромосомные перестройки могут происходить и в пределах одной хромосомы, и между хромосомами. Внутри хромосомы возможны потеря части хромосомы (делеция), удвоение участка (дупликация), перевертывание отдельных участков хромосомы на 180° (инверсия) и некоторые другие изменения. К межхромосомным перестройкам относят обмен участками между негомологичными хромосомами, который изменяет расположение генов в группах сцепления (транслокация). Хромосомные перестройки, подобно мутациям, могут приводить к различным уродствам или иметь летальный исход, но в то же время они являются механизмом, обеспечивающим дополнительный источник изменчивости в эволюции. Кроме перестроек в самих хромосомах, существуют хромосомные аномалии, связанные с нерасхождением хромосом в мейозе. Это приводит к тому, что в гамете может не хватать какой-либо хромосомы или иметься лишняя. Большинство таких аномалий приводит к гибели плода еще на ранних стадиях развития, однако некоторые нерасхождения хромосом обусловливают рождение аномальных индивидов, которые являются жизнеспособными. Наиболее известными из них являются люди с лишней 21-й хромосомой - больные с синдромом Дауна. Выживают также люди, которые имеют нехватку или избыток половых хромосом.
Выводы
Гены в хромосомах расположены линейно. Место в хромосоме, где расположен данный ген, называется локусом.
Различные состояния одного локуса носят название аллелей.
Явление, при котором данному локусу в популяции может соответствовать более двух аллелей, называется множественным аллелизмом.
Носитель одинаковых аллелей в гомологичных хромосомах называется гомозиготным. Носитель разных аллелей - гетерозиготным.
Ошибки, возникающие в процессе удвоения (репликации) ДНК, приводят к генным мутациям.
Генные мутации представляют собой изменения в последовательности нуклеотидов ДНК (замена, потеря или добавление).
Генные мутации, происходящие в гаметогенезе, то есть при образовании гамет, если они не летальны, увеличивают генетическое разнообразие, повышают количество гетерозигот, приводят к возникновению множественного аллелизма.
В процессе мейоза могут возникать перестройки хромосом, отличающиеся от нормальной рекомбинации (потеря участков, их удвоение, инверсия и др.), возможно также нерасхождение гомологичных хромосом во время редукционного деления.
3.5. Гены в популяциях. Закон Харди-Вайнберга
В теме
2 мы рассмотрели некоторые
особенности популяций
и основные типы признаков. В настоящей
теме мы познакомились с законами Менделя,
ввели понятие гена
и сформулировали основные положения
хромосомной теории наследственности.
Попробуем теперь совместить имеющиеся
у нас сведения и рассмотреть особенности
распространения не признаков, а генов
в популяциях. Когда мы анализировали
моно- и дигибридное скрещивание и
рисовали решетку
Пеннета, мы имели дело
с отдельной семьей и потомством от
единичного скрещивания. Теперь нашим
объектом будет популяция.
В
качестве удобной модели расщепления
по одной паре аллелей
может служить наследование групп крови
системы MN. Группа крови этой системы
определяется двумя аллелями М и N.
Гомозиготы
ММ имеют группу крови М, гомозиготы NN
имеют группу крови N, а гетерозиготы
MN имеют группу крови MN.
Рассмотрим
замкнутую ч еловеческую
популяцию, в которой имеется какое-то
количество аллелей М и какое-то количество
аллелей N. В принципе, частота встречаемости
аллелей М и N в популяции может меняться
от 100% М, когда вся популяция представлена
только гомозиготами ММ, до 100% N, когда
вся популяция представлена только
гомозиготами NN. Если аллели встречаются
с одинаковой частотой, то частота
встречаемости каждого из них составит
50%, или 0,5. Предположим, что в нашей
популяции представлены не только
гомозиготы одного типа, а все три типа
сочетаний аллелей, и частота встречаемости
каждого аллеля составляет 0,5. Понятно,
что в такой популяции с равной вероятностью
будут производиться гаметы,
несущие аллель М и аллель N, т.е. частоты
встречаемости этих гамет также будут
равны 0,5. Если браки в этой популяции
происходят случайно, то мы можем
воспользоваться решеткой Пеннета и
изобразить частоту образования гомозигот
и гетерозигот в следующем поколении
(вероятности перемножаются).
Мы
видим, что частоты встречаемости
гомозигот (ММ и NN) составляют по 0,25, тогда
как частота встречаемости гетерозигот
(MN) в два раза выше - 0,5. Частоты же
встречаемости каждого аллеля (М и N)
будут по-прежнему одинаковыми - по 0,5.
С
еловеческую
популяцию, в которой имеется какое-то
количество аллелей М и какое-то количество
аллелей N. В принципе, частота встречаемости
аллелей М и N в популяции может меняться
от 100% М, когда вся популяция представлена
только гомозиготами ММ, до 100% N, когда
вся популяция представлена только
гомозиготами NN. Если аллели встречаются
с одинаковой частотой, то частота
встречаемости каждого из них составит
50%, или 0,5. Предположим, что в нашей
популяции представлены не только
гомозиготы одного типа, а все три типа
сочетаний аллелей, и частота встречаемости
каждого аллеля составляет 0,5. Понятно,
что в такой популяции с равной вероятностью
будут производиться гаметы,
несущие аллель М и аллель N, т.е. частоты
встречаемости этих гамет также будут
равны 0,5. Если браки в этой популяции
происходят случайно, то мы можем
воспользоваться решеткой Пеннета и
изобразить частоту образования гомозигот
и гетерозигот в следующем поколении
(вероятности перемножаются).
Мы
видим, что частоты встречаемости
гомозигот (ММ и NN) составляют по 0,25, тогда
как частота встречаемости гетерозигот
(MN) в два раза выше - 0,5. Частоты же
встречаемости каждого аллеля (М и N)
будут по-прежнему одинаковыми - по 0,5.
С оответственно
в следующем поколении этой популяции
(F2), при сохранении случайности браков,
будут опять получены те же
соотношения.
В
реальных популяциях, как правило,
наблюдаются самые различные частоты
встречаемости аллелей, к тому же между
аллелями могут существовать отношения
доминантности
и рецессивности,
и гетерозиготы по внешней выраженности
признака могут совпадать с доминантным
типом гомозигот, т.е. частоты встречаемости
самого признака будут отличаться от
частот встречаемости гомозигот
и гетерозигот.
Разберем
еще один пример, связанный со вкусовой
чувствительностью. Когда мы говорили
о дискретной изменчивости, мы упоминали
о существовании двух типов людей, имеющих
разную чувствительность к вкусу
фенилтиомочевины
(ФТМ). Эта чувствительность зависит от
одного гена, представленного парой
аллелей. Доминантный аллель Т определяет
чувствительность к вкусу ФТМ, а рецессивный
аллель t - отсутствие чувствительности.
Таким образом, гомозиготы tt не ощущают
горького вкуса ФТМ, тогда как гомозиготы
ТТ и гетерозиготы Tt его ощущают. Опять
представим себе изолированную человеческую
популяцию, в которой браки осуществляются
случайно, а частоты встречаемости
аллелей Т и t составляют 0,6 и 0,4
соответственно. Естественно, что частоты
гамет, несущих данные аллели,
будут такими же, и мы опять можем
изобразить с помощью решетки Пеннета
частоту различных сочетаний аллелей в
поколении F1.
оответственно
в следующем поколении этой популяции
(F2), при сохранении случайности браков,
будут опять получены те же
соотношения.
В
реальных популяциях, как правило,
наблюдаются самые различные частоты
встречаемости аллелей, к тому же между
аллелями могут существовать отношения
доминантности
и рецессивности,
и гетерозиготы по внешней выраженности
признака могут совпадать с доминантным
типом гомозигот, т.е. частоты встречаемости
самого признака будут отличаться от
частот встречаемости гомозигот
и гетерозигот.
Разберем
еще один пример, связанный со вкусовой
чувствительностью. Когда мы говорили
о дискретной изменчивости, мы упоминали
о существовании двух типов людей, имеющих
разную чувствительность к вкусу
фенилтиомочевины
(ФТМ). Эта чувствительность зависит от
одного гена, представленного парой
аллелей. Доминантный аллель Т определяет
чувствительность к вкусу ФТМ, а рецессивный
аллель t - отсутствие чувствительности.
Таким образом, гомозиготы tt не ощущают
горького вкуса ФТМ, тогда как гомозиготы
ТТ и гетерозиготы Tt его ощущают. Опять
представим себе изолированную человеческую
популяцию, в которой браки осуществляются
случайно, а частоты встречаемости
аллелей Т и t составляют 0,6 и 0,4
соответственно. Естественно, что частоты
гамет, несущих данные аллели,
будут такими же, и мы опять можем
изобразить с помощью решетки Пеннета
частоту различных сочетаний аллелей в
поколении F1.
Таким образом, в поколении F1 частоты гомозигот и гетерозигот будут следующими:
ТТ = 0,62 = 0,36;
Tt = 2.0,4.0,6 = 0,48;
tt = 0,42 = 0,16.
Заметим, что в сумме это составляет единицу. Если рассмотреть частоту встречаемости самого признака, то окажется, что в данной популяции число людей, не ощущающих вкус ФТМ, составит 16%. Теперь посмотрим, каковы будут гаметы, производимые новым поколением. Гомозиготы ТТ производят лишь гаметы типа Т; гетерозиготы Tt с равной вероятностью производят гаметы обоих типов (Т и t); гомозиготы tt производят лишь гаметы типа t
Следовательно, частоты встречаемости гамет Т и t будут следующими:
Т = 0,36 + 1/2.0,48 = 0,6;
t = 0,16 + 1/2.0,48 = 0,4;
Отсюда ясно, что частоты аллелей при замкнутости популяции и случайности браков сохранились и в следующем поколении. Все это может быть записано в виде буквенных выражений. Если частоты аллелей обозначить буквами р и q, то мы можем записать, что в популяции р + q = 1. Решетку Пеннета для аллелей Т и t мы можем представить так:
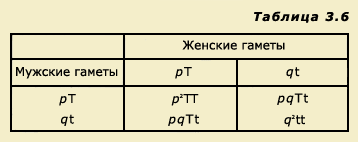
Таким образом, мы можем записать, что сумма частот встречаемости различных зигот равна:
р2 + 2pq + q2 = (р + q)2 .
Эта формула отражает распределение гомозигот и гетерозигот в популяции. Она была впервые предложена в 1908 г. двумя исследователями - математиком Г. Харди и врачом В. Вайнбергом, независимо друг от друга.
Выводы
3.1. Законы Менделя
Открытие дискретного характера наследственности принадлежит Г. Менделю.
Наследственные единицы передаются из поколения в поколение в неизменном виде.
Признаки организма детерминируются парами наследственных единиц.
При образовании половых клеток (гамет) парные наследственные единицы расходятся, и в каждой гамете бывает представлена лишь одна из них.
В процессе образования гамет наследственные единицы перегруппировываются (рекомбинируются), что приводит к новым сочетаниям признаков у потомства.
3.2. Хромосомная теория наследственности
Носителями единиц наследственности являются структуры клеточного ядра - хромосомы.
Хромосомы легко могут наблюдаться в делящихся клетках. В клетках тела содержится диплоидный (двойной) набор хромосом - каждая хромосома имеет аналогичную себе сестринскую хромосому (гомологичные хромосомы). В половых клетках содержится гаплоидный (одинарный) набор хромосом.
В клетках тела человека 46 хромосом (23 пары).
Существует два типа клеточного деления - митоз и мейоз. Первый характерен для деления соматических клеток, второй происходит при образовании половых клеток (гамет).
При митозе хромосомы удваиваются и затем расходятся по дочерним клеткам. В результате образуются две клетки, абсолютно идентичные родительской.
При мейозе хромосомы удваиваются один раз, но затем следуют два цикла клеточных делений. При первом делении (редукционном) гомологичные хромосомы случайным образом расходятся по разным клеткам. Второе деление мейоза напоминает митоз. В результате мейоза образуется четыре дочерних клетки с гаплоидным набором хромосом.
Процесс рекомбинации хромосом при редукционном делении соответствует рекомбинации менделевских единиц наследственности.
Единицы наследственности называются генами и располагаются линейно в хромосомах. Гены, расположенные в одной хромосоме, называются сцепленными.
Сцепленные гены могут рекомбинировать благодаря процессу кроссинговера, при котором происходит обмен участками между гомологичными хромосомами.
Процессы рекомбинации, происходящие в мейозе, лежат в основе генетической изменчивости и приводят к генетической уникальности индивидов.
3.3. Молекулярные основы наследственности
Материальным субстратом наследственности являются молекулы дезоксирибонуклеиновой кислоты (ДНК).
Молекулы ДНК способны к удвоению с большой точностью воспроизведения.
Молекулы ДНК способны образовывать бесконечное разнообразие различных форм.
ДНК представляет собой цепь нуклеотидов, в состав которых входят три компонента - фосфорный, углеводный и азотистое основание (аденин, гуанин, тимин или цитозин).
Молекула ДНК состоит из двух полинуклеотидных цепей, соединенных через азотистые основания, и имеет комплементарное строение: связи между нитями образуются только в парах аденин-тимии (А-Т) и гуанин-цитозин (Г-Ц).
Генетическая информация кодируется последовательностью оснований в цепи ДНК.
Основной функцией гена является кодирование информации для синтеза специфического белка.
Аминокислоты для синтеза белка кодируются триплетами оснований в цепи ДНК (генетический код).