

Глава yii фотосинтез источники углерода для растений. Сущность фотосинтеза
При рассмотрении химического состава растений отмечалось, что углерод составляет почти половину их сухого вещества. Атомы углерода образуют скелет всех органических соединений, а способность их вступать в реакцию с другими элементами обусловливает огромное число этих соединений.
Опыты с водными культурами с большой достоверностью показали, что подавляющую часть углерода растения получают не из почвы. С другой стороны, если поместить растение в атмосферу, очищенную от углекислого газа, например, под стеклянный колпак, в который воздух поступает только через слой натронной извести, то оно начнет обнаруживать признаки голодания. Это свидетельствует о том, что подавляющую часть потребности в углероде растения покрывают за счет углекислого газа атмосферы, составляющего 0,03% общего объёма воздуха.
Добывание растением из углекислого газа воздуха нужного им для построения своего тела углерода представляет один из важнейших
процессов в жизни растения и всего органического мира. Животные и чело век не могут усваивать углерод непосредственно из углекислого газа и для питания употребляют уже выработанные растением органические со- единения.
Организмы, использующие для питания углекислый газ воздуха, называются автотрофными. Организмы, не способные усваивать углекислый газ воздуха и питающиеся за счет органических веществ, выработанных другими живыми существами, называются гетеротрофными. Кроме животных к ним принадлежат незеленые растения — грибы, бактерии, некоторые высшие растения и др.
Для того чтобы растение могло использовать углекислый газ воздуха, необходимы совершенно определенные условия: свет и наличие хлорофилла. Процесс образования органических веществ из неорганических — углекислого газа и воды, происходящий в зеленых растениях на свету, называется фотосинтезом, или ассимиляцией. Его можно выразить следующим схематическим уравнением:

Фотосинтез — это окислительно-восстановительный процесс: от молекулы воды отнимается водород (окисление), который восстанавливает молекулу СО2. Исследования советских (А. Н. Виноградов, Р. В. Тейс) и американских (С. Рубен, М. Камен и др.) ученых показали, что свободный кислород выделяется из молекулы воды, а не из молекулы углекислого газа, как это считалось раньше. Своеобразие фотосинтеза заключается в том, что он, в отличие от подавляющего большинства других процессов, идет с увеличением свободной энергии системы. Поглощенная пигментами солнечная энергия не растрачивается, а накапливается в продуктах реакции в форме потенциальной химической энергии.
ЛИСТ КАК ОРГАН ФОТОСИНТЕЗА.
ХЛОРОПЛАСТЫ
Углекислый газ усваивается в зеленых пластидах клетки — хлоропластах. Следовательно, чтобы послужить материалом для синтеза углеводов, углекислый газ должен быть поглощен клетками, содержащими хлоропласты. Такие клетки составляют основную массу листа — мезофилл. Сверху лист покрыт эпидермой и кутикулой, мало проницаемой для газов. Основной путь, по которому углекислый газ проникает внутрь листа, — устьица. Хотя площадь устьичных отверстий, даже когда они полностью открыты, составляет незначительную часть всей листовой поверхности (не более 1%), диффузия газа через них, в соответствии с законами физики (закон Стефана), идёт с большой скоростью и эпидерма не представляет почти никакого препятствия для проникновения углекислого газа внутрь листа. Мезофилл листа обычно состоит из палисадной (столбчатой) и губчатой паренхимы. Палисадная паренхима расположена на верхней стороне листа и состоит из плотно прилегающих друг к другу клеток, вытянутых перпендикулярно к поверхности листа и богатых хлоропластами. Эту ткань можно рассматривать как ассимиляционную по преимуществу. Форма палисадных клеток способствует оттоку продуктов ассимиляции. Нижние концы клеток примыкают к особым собирающим клеткам, которые в свою очередь сообщаются с проводящими пучками. Благодаря этому устанавливается постоянный ток ассимилятов из листьев в другие органы растения. Губчатая паренхима находится ближе к нижней эпидерме. Клетки ее расположены рыхло и содержат значительно меньше хлоропластов. Считают, что эта ткань облегчает проветривание листа. Достигнув поверхности хлорофиллоносных клеток, углекислый газ растворяется в водe, которая всегда пропитывает их стенки. Затем уже в виде Н2СОз он диффундирует сквозь стенку, проникает в цитоплазму и достигает зеленых пластид, которыми поглощается.
Общая поверхность хлоропластов огромна. Так, в листе бука она примерно в 200 раз превышает его площадь и у столетнего дерева достигает 2 га. Это значительно облегчает растениям добывание углекислого газа из воздуха. Хлоропласты состоят из белково-липидной стромы и пигментов, легко извлекаемых органическими растворителями. Важнейший из пигментов — хлорофилл. Кроме хлорофилла, хлоропласты содержат желтые пигменты — каротиноиды (каротин и ксантофилл). Хлорофилл выполняет функцию поглощения световой энергии и переносит ее на восстановление углекислоты, а также химически участвует в этом процессе. Однако он функционирует только в сочетании со стромой. Строма является носителем ферментов, участвующих в сложных реакциях фотосинтеза. В строме из продукта фотосинтеза – сахара вырабатывается крахмал (первичный или ассимиляционный). Если затенить отдельные участки листа, то на светлом фоне получаются темные фигуры ( проба Сакса).


Рнс. . Обнаружение первичного крахмала при помощи пробы Сакса. А—лист, частично затененный; Б — лист после обработки спиртом и йодом
По химической природе хлорофилл — сложный эфир дикарбоновой кислоты — хлорофиллина и двух спиртов — метилового и фитола р. Хлорофилл содержит четыре соединенных между собой остатка пиррола, которые образуют порфириновое кольцо, центральный атом которого — Mg. По строению хлорофилл весьма близок к красящему веществу крови — гему. В его состав также входит порфириновое кольцо, однако в центре расположен атом Fe. Это сходство было показано Ч. В. Ненцким и польским ученым Л. Мархлевским. К. А. Тимирязев считал установление этого сходства едва ли не самым крупным открытием в области химического изучения хлорофилла.
Исследование многих сотен самых разнообразных видов высших растений показало, что хлорофилл у них совершенно одинаковый. Общее количество хлорофилла в растениях составляет около 1 % от сухого веса. Хлорофилл в хлоропластах находится не в свободном виде, а связан с белком, образуя хлороглобин.
Для образования хлорофилла в растениях необходимо несколько совершенно определенных условий: присутствие пропластид, способных к позеленению, света и солей железа.
Развивающиеся в темноте растения имеют желтый цвет. Они называются этиолированными. Если их выставить на свет, они быстро зеленеют. Считают, что они содержат особое вещество, называемое протохлорофиллом,_образуюшееся в.темноте_и_ под влиянием света_легко_ превращающееся в хлорофилл.
Если выращивать растения при полном отсутствии солей железа, то они тоже будут бледно-желтого цвета и быстро гибнут от истощения. Это явление называется хлорозом. Поскольку железо не входит в состав хлорофилла, считают, что оно служит специфическим катализатором, без которого не осуществляются какие-то подготовительные стадии позеленения. Хлороз часто наблюдается в природе, особенно у растений, растущих на почвах, богатых известью.
Кроме того, у растений иногда наблюдается явление альбинизма — неспособность образовывать хлорофилл даже при самых благоприятных для этого условиях.
УЧАСТИЕ ПИГМЕНТОВ В ПОГЛОЩЕНИИ СВЕТА. РАБОТЫ К. А. ТИМИРЯЗЕВА
Хлорофилл обладает избирательным поглощением световой энергии. Наиболее интенсивно поглощение происходит в красных лучах спектра (длина волны от 650 до 680 ммк) и сине-фиолетовых (длина волны около 470 ммк). Зеленые лучи и часть красных не поглощаются, они и придают хлорофиллу изумрудно-зеленый цвет. Желтые пигменты — каротин и ксантофилл — поглощают свет в зеленой и синей частях спектра.
Энергетическая сторона процесса фотосинтеза глубоко вскрыта и разъяснена в работах К. А. Тимирязева. Он показал, что фотосинтез осуществляется только в лучах спектра, поглощаемых хлорофиллом. Дальнейшие исследования полностью подтвердили это положение. Процесс фотосинтеза в различных частях спектра идет неодинаково. К. А. Тимирязев показал, что максимум ассимиляции приходится на красные лучи, которые несут максимум энергии и полнее всего поглощаются хлорофиллом. В сине-фиолетовых лучах ассимиляция идет слабее, так как они несут меньше энергии. Вопросу о значении в фотосинтезе отдельных частей спектра К. А. Тимирязев придавал большое принципиальное значение. До него господствовало мнение, что свет служит только раздражителем. Этой точки зрения придерживались и современники К. А. Тимирязева немецкие ученые Ю. Сакс и В. Пфеффер. К.А. Тимирязев показал, что свет является источником энергии и необходим для фотосинтеза.
На возбуждение одной молекулы хлорофилла требуется один квант, поэтому в красных лучах, несущих большое число мелких квантов, большее число его молекул перейдет в возбужденное состояние
Кроме избирательного поглощения световой энергии, хлорофилл обладает свойством флуоресценции: в отраженном свете он кажется кроваво-красным, так как отражает поглощенные лучи с изменением длины их волны. Это указывает на значительную фотохимическую активность хлорофилла. Коэффициент поглощенной лучистой энергии в фотосинтезе чрезвычайно низок- 1%- 5%, редко до 10%. Большая часть переходит в тепловую энергию, либо повышает температуру и рассеивается в окружающем пространстве.
ХИМИЗМ ФОТОСИНТЕЗА
Несмотря на простоту суммарного уравнения фотосинтеза, этот процесс отличается чрезвычайной сложностью. Это обусловливается сложностью углеводной молекулы, которая не может сразу возникнуть из таких простых веществ, как СО2 и Н2О; трудностью окисления и восстановления этих прочных соединений; участием в реакциях световой энергии. Исследования показали, что фотосинтез включает не только несколько фотохимических реакций, но и ряд ферментативных, так называемых темновых реакций.
Использование методов меченых атомов (изотопов С, Р, О, N), разделительной хроматографии на бумаге, электрофореза, ионообменной

Рис. 134. Цикл фотосинтетических превращений углерода по Кальвину
очистки и разделения и некоторых других позволило выявить химизм фотосинтеза.
Рядом работ установлено, что первым этапом усвоения углекислоты является присоединение СО2 к какому-то акцептору (веществу, воспри- нимающему, присоединяющему другое вещество), карбоксилирова-
ние его:
R • Н + СО2_→ R • СООН.
Таким образом, фотосинтетическому восстановлению подвергается углерод не углекислого газа, а карбоксильной группы. Широкие исследования по выяснению природы первичных акцепторов и путей фотосинтетического превращения углерода были проведены американским ученым Кальвином и его сотрудниками. Схема фотосинтетических превращений углерода, по Кальвину, представлена на рис. 134. Он считает, что процесс фотосинтеза носит циклический и разветвленный характер: одна ветвь этого цикла ведет к образованию прямых устойчивых продуктов фотосинтеза — углеводов, другая имеет циклический характер и приводит к образованию акцептора СО2 — рибулезодифосфата, который вовлекает в фотосинтетический цикл все новые и новые молекулы СО2.
Наряду с этими сложными превращениями углерода за счет энергии света образуются богатые энергией органические соединения фосфора, в . частности аденозинтрифосфат (АТФ). Этот процесс называется фотосинтетическим фосфорилированием:

Аденозинтрифосфорная кислота (АТФ)
Энергия макроэргических (богатых энергией, ~) фосфатных связей АТФ идет на восстановительные процессы. При гидролизе макроэргических связей освобождается 7000—16 000 кал на грамм-молекулу.отщепленного фосфата.
К световым реакциям фотосинтеза относится:
1) разложение воды (активированный хлорофилл + 2Н2О-инактивированный хлорофилл + +4Н+2О),
2) фотосинтетическое фосфорилирование,
3) синтез аминокислот и белков.
К темповым реакциям фотосинтеза относятся:
1) фиксация СО2 акцептором,
2) перенос активного водорода на соединение, в котором фиксирована молекула СО2,
3) восстановление акцептора СО2,
4) образование Сахаров.
Приведенное описание фотосинтетического превращения углерода далеко не исчерпывает всей сложности этого процесса. В частности, считают, что акцептором СО2 может быть не только рибулезодифосфат, но и другие соединения.
-
МИНЕРАЛЬНОЕ ПИТАНИЕ РАСТЕНИЙ
ВЕЩЕСТВА, ПОЛУЧАЕМЫЕ РАСТЕНИЯМИ ИЗ ПОЧВЫ
Нет ни одного элемента, который бы не был обнаружен в растениях. Элемент может быть случайной примесью и накапливаться в растениях в больших количествах, или же находиться в нем в ничтожно малом количестве, но безусловно необходим. Установлено, что растение может успешно развиваться, если в питательном растворе находится лишь семь элементов: К, Са, Mg, S, Fe, N и Р. Это мнение держалось в науке более 50 лет, но было установлено, что многие другие элементы играют важную роль в жизни растений. Было установлено, что для нормального роста и развития растений необходимы в ничтожно малых количествах Mg, Zn, Cu, Al, I, Md и др.
Получаемые растениями из почвы вещества по химической природе можно разделить на две группы: металлоиды и металлы.
Металлоиды поступают в растения в виде анионов соответствующих солей. Они необходимы для образования органических веществ. Металлы поступают в растения в виде катионов. Они находятся в клетках в свободном состоянии или слабосвязанном и служат регуляторами жизненных процессов. Например, магний входит в состав хлорофилла, железо и медь- ферментов и т, д.
МЕТАЛЛОИДЫ
Азот. Поступает в растение в виде анионов NO3 и NO2, так и виде катиона NН4. Его значение в жизни растений очень велико.
Фосфор воспринимается растениями в виде анионов солей фосфорной кислоты РО4 .В белковые молекулы он входит в той же окисленной форме. В растениях образуются сложные эфиры фосфорной кислоты и фосфатиды которые являются необходимой составляющей цитоплазмы как и белки. Фосфор стоит в центре всего энергетического обмена клетки. Витамины и некоторые ферменты проявляют свое действие лишь в соединении с фосфорной кислотой.
При разложении растительных остатков фосфорная кислота высвобождается в виде неорганических солей и может снова использоваться растениями.
Сера усваивается лишь в виде аниона серной кислоты SO4, источником ее служат растворимые соли. Используется для синтеза белка за счет продуктов фотосинтеза- углеводов, входит в состав горчичных и чесночных масел, участвует в дыхании и росте.
При перегнивании растительных остатков сера отщепляется от белковой молекулы в виде сероводорода, который не усваивается растениями и очень ядовит для корней. В пригодную форму он переводится серобактериями, окисляющими сероводород и серную кислоту.
МЕТАЛЛЫ
Калий содержится в клетках меристемы и молодых органах. Много калия в корнеплодах, клубнях, крахмалистых семенах. Калий обладает большой подвижностью. Из старых, отмирающих органов передвигается в более молодые жизнедеятельные части растения( реутилизация). Калийные удобрения почти всегда оказывают благоприятное воздействие на урожайность сельскохозяйственных культур.
Натрий присутствует в золе растений часто в больших количествах, однако особого значения для жизнедеятельности не имеет и может быть исключен из питательного раствора. Только голофиты- растения, свойственные засоленным почвам, лучше растут в присутствии солей натрия. Из культурных растений к этому типу относятся сахарная свекла, дикий предок которой произрастает на засоленных почвах Средиземного моря.
Магний содержится преимущественно в молодых органах и семенах ( до 10-15% золы). Физиологическое действие его близко к действию калия. Магний входит в состав некоторых металлоорганических соединений, в частности хлорофилла , он может активизировать действие некоторых ферментов . Влияние магния зависит от состава почвы. Резко отзываются на внесение магнезиальных удобрений растения, растущие на легких песчаных и супесчаных почвах.
Кальций необходим для роста молодых тканей. Он входит в состав цитоплазматических структур и ядер. Соединения кальция с пектиновыми веществами составляют основу срединных пластинок, склеивающих стенки клеток друг с другом. Многие ферменты активны лишь в присутствии ионов кальция. Он способствует увеличению вязкости цитоплазмы и влияет на поступление веществ в клетку. Одна из важных функций кальция – нейтрализация щавелевой кислоты, образующейся как побочный продукт обмена веществ. При отсутствии кальция наблюдается резкое угнетение корневой системы, Особенно много кальция в старых органах.
МИКРОЭЛЕМЕНТЫ
Микроэлементы необходимы растениям в ничтожно малых количествах и в больших дозах становятся ядовитыми.
Железо участвует в образовании хлорофилла как катализатор. Оно входит в состав окисленных ферментов, играет чрезвычайную роль в процессе дыхания. Возможно железо участвует в процессе фотосинтеза и окислительно-востановительных процессах в клетке как переносчик электронов.
Цинк входит в состав некоторых ферментов. При отсутствии цинка наблюдается угнетение роста молодых проростков, заболевание цитрусовых и тунга.
Марганец активизирует работу многих ферментов, играет большую роль в восстановлении нитратов в растении, влияет на окислительно-востановительные процессы превращения железа. Для нормального роста растениям необходимо ничтожно малое количества марганца, поэтому внесение марганцевых удобрений не всегда дает положительный результат. Отзывчивы на такие удобрения сахарная свекла, хлопчатник, табак и другие культуры.
Бор при его недостатке у растений отмирают точки роста., нарушается расположение элементов ксилемы и флоэмы и полная потеря ими проводимости. Бор благоприятно влияет на цветение и плодоношение. Борные удобрения необходимы на известковый почвах. Очень отзывчивы на эти удобрения сахарная свекла, бобовые травы. При повышенных концентрациях он оказывает угнетающее действие на растения.
Медь оказывает влияние на окислительно-востановительную систему, входит в состав ряда ферментных систем.
При недостатке того или иного элемента у растений наблюдаются определенные симптомы повреждений, по которым можно определить, какого элемента не хватает. Так, хлороз листьев показывает на нехватку железа, отмирание корней- недостаток кальция, отмирание точек роста на недостаток бора. Однако такая диагностика улавливает далеко зашедшую форму голодания, которую внесением удобрений уже не исправить.
ПЕРЕДВИЖЕНИЕ ВЕЩЕСТВ В РАСТЕНИЯХ.
ВОДНЫЙ РЕЖИМ РАСТЕНИЙ. ДЕЙСТВИЕ НА РАСТЕНИЯ ВЫСОКИХ И НИЗКИХ ТЕМПЕРАТУР
ДВА ТОКА ВЕЩЕСТВ В РАСТЕНИИ
Существование организма растения как единого целого, физиологическая взаимосвязь отдельных органов, расположенных в неодинаковых физических средах и выполняющих различные функции, возможны лишь при условии передвижения минеральных и органических веществ.
Факт передвижения веществ в растении по двум направлениям был установлен еще в 1679 г. Мальпиги путем кольцевания. Если удалить со стебля участок коры в виде кольца, то листья на нем остаются живыми и не обнаруживают никаких признаков завядания, а плоды бывают даже больше, чем на не окольцованных ветвях. Это показывает, что передвижение воды и минеральных веществ из почвы осуществляется по древесине (ксилеме). Этот ток веществ был назван восходящим. У верхнего же края кольцевого выреза происходит застой питательных веществ и разрастание тканей коры в виде наплыва. Если наплыв не восстановит удаленную часть коры, то корни отмирают от истощения и все растение погибает. Значит, пластические вещества из листьев в корень передвигаются по коре (в основном по флоэме). Этот ток веществ был назван нисходящим.
Очень долго в науке держалось мнение, что по флоэме передвигаются только органические вещества, а по ксилеме — только вода и минеральные вещества. Однако исследования последних лет с применением метода меченых атомов показали, что по флоэме могут передвигаться не только органические, но и минеральные вещества. В основном это калий, фосфор, частично кальций. Причем эта миграция может осуществляться в любом направлении. После того как было доказано, что в корне тоже синтезируются органические вещества, стало ясно, что и по ксилеме вверх передвигаются не только минеральные, но и органические вещества. Кроме того, установлено, что минеральные и органические вещества из корня могут подниматься вверх по флоэме.
Законы, управляющие передвижением органических веществ, до сих пор изучены мало. Исследования показали, что скорость передвижения органических веществ во много раз больше скорости диффузии, что проводящие пучки отличаются очень интенсивным дыханием и что клетки флоэмы способны не только проводить органические вещества, но и подвергать их различным превращениям. Это позволило сделать предположение, что органические соединения перемещаются по флоэме не вследствие пассивного перетекания растворов или диффузии, а в результате каких-то обменных реакций, непрерывно происходящих в ситовидных трубках между цитоплазмой и передвигающимися молекулами.
ПЕРЕДВИЖЕНИЕ ВОДЫ В РАСТЕНИИ
Путь, который проходит вода в растении, делится на две части: 1) по живым клеткам от корневого волоска до сосудов центрального цилиндра корня и от сосудов листа до клеток мезофилла, испаряющих воду в межклетники; 2) по мертвым клеткам проводящей системы — сосудам и трахеидам.
Путь воды по живым клеткам исчисляется миллиметрами, однако он представляет большие трудности, так как при переходе от одной клетки к другой вода встречает значительное сопротивление, поэтому этим способом вода не может передаваться на большие расстояния. Большую часть пути вода проходит по мертвым, пустым, вытянутым в длину клеткам — трахеидам или же по полым трубкам — сосудам.
Поглощение воды и ее перемещение вверх осуществляется в результате совместного действия следующих факторов: корневого давления (нижний концевой двигатель), транспирации (верхний концевой двигатель), сил сцепления молекул воды.
. Жидкость, вытекающая при плаче растений, называется пасокой. Химический состав ее непостоянен. Весной, когда происходит гидролиз запасных углеводов, она богата сахарами, органическими кислотами и содержит мало минеральных веществ. Выделение капельно-жидкой воды может происходить и через листья, через особые водяные устьица — гидатоды Это явление получило название гуттации. Гуттация происходит в умеренно теплой и влажной, насыщенной водяными парами атмосфере, когда возникает диспропорция между поступлением воды и ее испарением. Наиболее часто она встречается у растений тропической и субтропической зон и иногда происходит с такой силой, что создает впечатление дождя. Из растений умеренной зоны активно гуттируют ива, картофель, гречиха и др. Плач растений и гуттация не являются только осмотическими процессами, так как они прекращаются при действии на корни веществами, ингибирующими дыхание. Прежде чем попасть в сосуд корня, вода, поглощенная корневым волоском, должна проделать путь по живым клеткам коровой паренхимы. Согласно Д. А. Сабинину, такой односторонний ток воды возможен только при различии в обмене веществ в разных частях клетки, при котором на одном полюсе клетки образуется больше тургорогенных веществ, чем на другом, а следовательно, возникают большее осмотическое давление и большая сосущая сила. Поступление воды из клеток в сосуды происходит вследствие того, что раствор, находящийся в сосудах, обладает большей сосущей силой, чем близлежащие клетки. Сосуды — это мертвые клетки без цитоплазмы и их сосущая сила равна всей величине осмотического давления раствора (S = P), в то время как в живых клетках существует еще тургорное давление и S = P—Т. Вода, находящаяся в сосудах и трахеидах, имеет вид тонких нитей, которые своими нижними концами упираются в паренхимные клетки корня, а верхними — как бы подвешены к испаряющим клеткам листа. Для того чтобы вода передвигалась вверх, необходимо, чтобы испаряющие клетки обладали достаточной сосущей силой, которая тем больше, чем сильнее испарение. В клетках листьев древесных растений она достигает 10— 15 атм.
Однако, как показал русский ученый Е. Ф. Вотчал, поднятие воды на большую высоту по сосудам возможно лишь при условии существования непрерывных водных нитей, которое обеспечивается силами сцепления молекул воды между собой и со стенками сосудов. Сила сцепления достигает 300—350 атм.
ТРАНСПИРАЦИЯ
Испарение воды растением не только чисто физический, но и физиологический процесс, так как на него большое влияние оказывают анатомические и физиологические особенности растения. Этот процесс назван тринспирацией.
Испарение
воды в листе происходит с поверхности
клеток мезофилла, По вычислениям
Тэррела, эта поверхность у растений
умеренно влажных местообитаний в
12—19 раз превышает наружную поверхность
листьев, а у растений засушливых
местообитаний — в 17—30 раз. Парообразная
вода попадает в межклетники и через
устьичные щели диффундирует наружу.
Такая транспирация называется устьичной.
Площадь устьичных щелей составляет
около 1 % общей площади листа. Однако,
как уже отмечалось в отношении диффузии
газа при фотосинтезе, диффузия пара
через устьица идет с той же скоростью,
с какой она шла бы при отсутствии
эпидермы. Одна из важнейших особенностей
устьичного аппарата — способность
открывать и закрывать устьичные
отверстия. Стенка замыкающих клеток
имеет неодинаковую толщину: часть
стенки, примыкающая к щели, значительно
утолщена, в то время как остальная часть
стенки остается тонкой. Это приводит
к тому, что при насасывании воды тонкая
наружная часть стенки растягивается
значительно больше, чем толстая,
кривизна клеток увеличивается, и щель
раскрывается. При уменьшении объема
замыкающей клетки тонкая стенка
распрямляется, и щель закрывается. В
основе процесса, обусловливающего
изменение тургора в замыкающих клетках,
лежит превращение крахмала в сахар и
обратно, которое вызывается изменением
хода ферментативных реакций. Большое
влияние на открывание устьиц

Рис. 143. Изменение транспирации в зависимости от изменения основных метеорологических показателей в течение дня:
/ — обшая солнечная радиация, 2 — дефицит насыщения, 3 — температура, 4 — интенсивность
транспирации
оказывает свет. На свету устьица закрываются лишь с большим трудом. Это фотоактивное открывание устьиц носит приспособительный характер: через устьица проникает в лист углекислый газ и для процесса фотосинтеза необходимо, чтобы устьица были открыты в светлые часы суток. Пользуясь различными методами, можно проследить за ходом устьичных движений в течение суток. В ясную не очень жаркую и сухую погоду у большинства растений устьичные щели открываются на рассвете, наиболее широко они раскрыты в утренние часы, к полудню они начинают сужаться и закрываются несколько раньше захода солнца. В сухую и жаркую погоду устьичные щели к полудню закрываются полностью, а к вечеру опять открываются. У разных растений устьица ведут себя неодинаково. Так, у картофеля, капусты и некоторых других растений устьица обычно открыты круглые сутки, у хлебных злаков устьица на ночь закрываются. Большинство растений в этом отношении занимает промежуточное положение. Движение устьичного аппарата в зависимости от внешних условий весьма сложны и не всегда поддаются учету.
Наряду с устьицами в испарении воды участвует и вся поверхность листьев, несмотря на то, что она покрыта кутикулой. Эта форма транспирации называется кутикулярной. У взрослых листьев кутикулярная транспирация в 10—20 раз слабее, чем устьичная.
Транспирация имеет следующее значение: 1) создает непрерывный ток воды, 2) облегчает передвижение минеральных веществ от корня к листьям, 3) защищает листья от перегрева.
Количество воды, которое растение пропускает через себя, огромно. Одно растение подсолнечника или кукурузы за вегетационный период испаряет более 200 кг воды.
Транспирация зависит от метеорологических условий: температуры воздуха, света, ветра, дефицита насыщения воздуха парами воды, а также от количества воды в растении. В результате получаются те сложные кривые, которые характеризуют суточный ход этого процесса в природе (рис. 143). Опыты показали, что для нормального развития растения не нуждаются в том огромном количестве воды, которое они теряют в естественных условиях и что транспирация очень часто может быть сокращена с пользой для них. Так, наиболее пышное развитие растений наблюдается во влажном тропическом климате, где влажность почвы и воздуха очень высоки. В оранжереях растения растут лучше, если поддерживать влажность на возможно более высоком уровне. Даже в полевой культуре применяют освежительные поливы дождеванием в целях повышения влажности воздуха и снижения транспирации.
ВОДНЫЙ БАЛАНС РАСТЕНИЙ
Растения, живущие на суше, должны поддерживать цитоплазму клеток в достаточно насыщенном водой состоянии. Поэтому они имеют ряд особенностей в своем строении, которые, с одной стороны, обеспечивают снижение количества теряемой воды (кутикула, покрывающая все надземные части, восковой налет, волоски и т. д.), а с другой — быструю подачу воды из почвы к листьям (мощная корневая система, хорошо развитая проводящая система и т. д.). В то же время, чтобы успешно шел процесс фотосинтеза, необходим тесный контакт хлорофиллоносных клеток с окружающей атмосферой. Это приводит к непрерывному испарению воды клетками, которое усиливается нагреванием листа вследствие поглощения хлорофиллом солнечной энергии, также необходимой для фотосинтеза. Это глубокое внутреннее противоречие между углеродным питанием и водным режимом К. А. Тимирязев назвал «необходимым злом», так как в условиях засухи оно может привести к гибели растения. Это противоречие кладет глубокий отпечаток на строение растений и на всю их жизнедеятельность.
Одно из важнейших условий нормального функционирования высших наземных растений — сведение водного баланса, т. е. соотношения между приходом и расходом воды, без длительного и глубокого дефицита. В умеренно влажные и не слишком жаркие дни это условие выдерживается. Но в ясные летние дни к полудню транспирация настолько увеличивается, что возникает водный дефицит, который при достаточной влажности почвы достигает 5—10%, а при недостатке влаги в почве возрастает до 25% и более. Это вполне нормальное явление. Дальнейшему увеличению водного дефицита препятствует способность растений под влиянием потери воды регулировать свою транспирацию в довольно широких пределах.
Однако эта регуляция имеет свои границы и при значительном возрастании транспирации и иссушении почвы наступает резкое нарушение водного баланса, которое внешне выражается в завядании. При этом клетки теряют тургор, листья, и молодые побеги повисают вниз. Завядаиие еще не означает утрату растением жизнедеятельности. Если растение своевременно снабдить водой, то тургор восстанавливается. Различают два типа завядания растений: временное и длительное. Первое наблюдается при сильном возрастании транспирации, когда поступающая из почвы вода не успевает покрывать ее трату. При этом листья, больше всего расходующие воду, теряют тургор и вянут, а остальные органы растения содержат еще достаточное количество воды. При ослаблении транспирации, например к вечеру, водный дефицит исчезает и растение оправляется без добавочного увлажнения почвы. Большого вреда растениям временное завядание не приносит, но все же снижает урожай, так как приостанавливает фотосинтез и рост. Длительное завядание наблюдается, когда почва не содержит достаточного количества доступной для растений воды. При этом водный дефицит за ночь не исчезает, и к утру: растения оказываются не вполне насыщенными водой и не в состоянии нормально функционировать. В этих условиях тургор постепенно падает во всех органах растения, вплоть до корневых волосков, так как завядшие листья, обладая большой сосущей силой, оттягивают от них воду. Корневые волоски отмирают, поэтому даже при обильном поливе растения восстанавливают прежнюю скорость водоснабжения только через несколько дней, когда образуются новые корневые волоски. Исследования Н. А. Максимова, Н. М. Сисакяна и других показали, что завядание вызывает глубокое влияние на состояние биоколлоидов клетки, которое ведет к нарушению обмена веществ. Процессы гидролиза усиливаются, синтетические процессы задерживаются. Это отражается на всех физиологических функциях растения — фотосинтезе, дыхании, передвижении веществ, росте и т. д. Урожай падает, зерно получается щуплым. Длительное завядание вызывает необратимые изменения, и клетки в конце концов отмирают даже при возобновлении водоснабжения. В то же время завядание — весьма действенный способ задержать транспирацию в наиболее опасные для растения периоды. В завядшем состоянии потеря воды растением в 5—10 раз меньше, чем в благоприятном периоде.
У разных растений завядание наступает при потере неодинакового количества воды. Так, подсолнечник и картофель не завядают при потере 25—30% воды, а другие растения, особенно теневые, завядают уже при потере 2—3% воды. Соотношение между добыванием и расходом воды зависит от многих факторов. Это обусловливает чрезвычайное разнообразие типов наземных растений по отношению к водному режиму.
ДЕЙСТВИЕ НА РАСТЕНИЯ НЕДОСТАТКА ВЛАГИ
И ВЫСОКИХ ТЕМПЕРАТУР.
ЗАСУХОУСТОЙЧИВОСТЬ И ЖАРОСТОЙКОСТЬ
Засуха — это резкое проявление недостатка влаги, приводящее к нарушению водного режима растений. Засуха бывает атмосферная и почвенная. Атмосферная засуха характеризуется высокой температурой и малой относительной влажностью воздуха (10—20%). Она приводит к завяданию растений. Сопровождающая атмосферную засуху высокая температура воздуха вызывает сильный нагрев растений. Большие повреждения растениям наносят суховеи — очень сухие горячие ветры. При этом высыхает и отмирает значительная часть листьев. При атмосферной засухе корневая система остается неповрежденной. При большой продолжительности атмосферная засуха вызывает иссушение почвы — почвенную засуху. Она более опасна для растения, так как приводит к длительному завяданию. Уже отмечалось, что завядание растений нарушает обмен веществ и значительно снижает урожайность.
Разные части растений реагируют на засуху неодинаково. Так, уменьшение содержания воды в листьях приводит к повышению их сосущей силы, и они начинают отсасывать воду от конусов нарастания стебля, бутонов и завязавшихся плодов. Это вызывает гибель цветков или их стерильность, образование щуплого зерна — захват. Верхние листья дольше сохраняют свою жизнедеятельность при засухе, чем нижние, так как оттягивают от них воду. Эта особенность верхних листьев объясняется тем, что они находятся в условиях несколько затрудненного водоснабжения и в связи с этим имеют ряд анатомических и физиологических признаков, свойственных растениям засушливых местообитаний — ксерофитам (ксероморфизм): мелкие клетки, большое число устьиц, густая сеть проводящих пучков, интенсивные фотосинтез и транспирация, высокая концентрация клеточного сока и т. д
В различные периоды онтогенеза растения неодинаково устойчивы к засухе. Наиболее чувствительным является период формирования репродуктивных органов и оплодотворения; он получил название критического.
Засухоустойчивость у растений выражается в способности сохранять нормальный или близкий к нему ход процессов обмена веществ в условиях водного дефицита, который у других форм растений вызывает необратимые патологические нарушения. Культурные растения подвергаются засухе обычно не весь вегетационный период, а в течение более или менее коротких промежутков времени. Поэтому важнейшая особенность засухоустойчивых сортов — способность переносить временное обезвоживание своих тканей без снижения урожая. Засухоустойчивость может быть повышена применением удобрений. Наиболее радикальная мера борьбы с засухой — искусственное орошение.
. Отмирание начинается уже при температуре немного выше 40° С, когда до свертывания белков еще далеко. При этом нарушается согласованность между идущими в клетках биохимическими процессами и образуются ядовитые вещества, отравляющие цитоплазму. Нарушается соотношение между фотосинтезом и дыханием: при повышенных температурах расход органических веществ на дыхание превышает их синтез, и растение начинает голодать. Высокие температуры разрушают также белково-липидный комплекс мембран цитоплазмы, что влечет за собой потерю клеткой осмотических свойств. При температурах выше 50°С к отравлению присоединяется еще и свертывание цитоплазмы, что ведет к быстрому отмиранию клеток.
В различные периоды индивидуального развития растения в неодинаковой степени чувствительны к повышенным температурам. Наиболее вредны они в ранние периоды роста и во время формирования цветков. При созревании плодов высокие температуры полезны. Важное условие защиты листьев от перегрева — беспрепятственная транспирация, поэтому сухая жара переносится растениями легче, чем влажная.
Перегрев вызывает повреждение растений, называемое запалом. Он обнаруживается в виде различно окрашенных некротических пятен на листьях, а также в виде щуплости зерна. В природе чаще наблюдается одновременное повреждение растений от перегрева и обезвоживания, поэтому порой трудно точно разграничить непосредственное влияние высокой температуры и засухи.
Наибольшей жаростойкостью — выносливостью к высокой температуре отличаются ксерофиты — растения, произрастающие в засушливых условиях. Из культурных растений наивысшая жаростойкость свойственна растениям южных широт (сорго, рис, хлопчатник, клещевина и др.). Устойчивость растений к перегреву обусловлена высокой вязкостью цитоплазмы их клеток и большим количеством воды, связанной ее коллоидами..Поэтому при нарастании водного дефицита, жаростойкость растений повышается и чем интенсивнее белковый обмен, тем легче растение переносит высокие температуры. Рост семени обычно заканчивается незадолго до завершения его полного физиологического развития. Несколько позднее прекращается приток питательных веществ и снижается активность растительных гормонов. По мере того как активность гормонов и ферментов снижается до минимума, влажность семян падает. Покровы семени претерпевают существенные изменения: их ткани частично отмирают, уплотняются и нередко одревесневают. Такие зрелые семена способны переносить неблагоприятные условия среды и могут длительно (иногда до нескольких десятков лет) сохранять способность к прорастанию. Такие зрелые семена находятся в физиологическом покое, в этом состоянии происходят метаболические процессы, дыхание, иногда "дозревание" зародыша, но способность к набуханию при поступлении влаги и прорастанию часто заторможена.
Физиология прорастания семян.
Степень глубины физиологического покоя и его длительность неодинаковы. Семена выводятся из состояния покоя различным образом. Некоторые семена, особенно однолетних растений, легко набухают и прорастают уже под влиянием увлажнения. Для прорастания других и нормального развития проростка обязательна холодная стратификация , т.е. длительное выдерживание их при пониженной температуре, во влажной среде и в условиях хорошей аэрации. Наконец, существует еще одна группа так называемых "твердосеменных", семенная кожура которых в силу ее структурных особенностей водонепроницаема. Такие семена прорастают только после скарификации - искусственного нарушения целостности кожуры с помощью надцарапывания, перетирания с песком, ошпаривания кипятком и т.д. В природе такие семена набухают и прорастают обычно под влиянием резкой смены температурных режимов, способствующих нарушению целостности оболочки.
Прорастанием семян называют их переход от состояния покоя к вегетативному росту зародыша и формированию из него проростка. Прорастание начинается при оптимальном для каждого вида сочетании влажности и температуры среды, при свободном доступе кислорода. Прорастание семян сопровождается сложными биохимическими и морфофизиологическими процессами. При поступлении воды в семенах резко усиливается процесс дыхания, активизируются ферменты, запасные вещества переходят в легкоусвояемую, подвижную форму, образуются полирибосомы и начинается синтез белка и других веществ.
Рост зародыша обычно начинается с прорыва покровов удлиняющимися зародышевым корнем и гипокотилем в области микропилярного следа . После появления корня почечка развивается в побег, на котором развертываются настоящие листья ( рис. 51 ).
В практике сельского хозяйства жизнеспособность и качество семян характеризуются всхожестью, т.е. процентом семян, давших нормальные проростки в оптимальных для них условиях за определенный срок. Для полевых культур этот срок равен 6-10 суткам, для древесных - до 2 месяцев.
-
НИЗШИЕ РАСТЕНИЯ —THALLOBIONTA
Большинство низших растений настолько обособлено вследствие длительного развития и специализации, что часто трудно определить общего предка. По числу видов и по распространению в самых разнообразных эколого-географических условиях низшие стоят на первом месте. Они состоят из двух резко различных по способу питания групп, первую из которых составляют автотрофные растения, в основном водоросли и лишайники, вторую — гетеротрофные: большинство бактерий, слизевики, грибы.
Низшие растения имеют выдающееся значение в природе. Для человека водоросли, грибы и лишайники представляют богатый резерв пищи, лекарственного и технического сырья. С другой стороны, многие бактерии, грибы и слизевики — злостные паразиты растений, животных и человека.
ОТДЕЛ БАКТЕРИИ — BACTERIOPHYTA
Бактерии — мельчайшие организмы, которые изучают с помощью особых методов. Наряду с мельчайшими животными организмами они являются объектом особой науки — микробиологии. Отдел охватывает около 3 тыс. видов.
Морфология. Важнейшие структурные особенности бактериальной клетки заключаются в отсутствии у большинства сформированного ядра (при значительном содержании нуклеиновых кислот), пластид и красящего вещества. Лишь изредка встречаются окрашенные автотрофные бактерии — пурпурные или зеленые. Клеточная стенка бактерий состоит из азотистых веществ, а не из целлюлозы, и способна к ослизнению.
В подавляющем большинстве бактерии очень малы по величине. Так, Bacillus prodigiosus, часто развивающийся на вареном картофеле, хлебе, мясе, достигает всего лишь 0,0008 мм (0,8 мк), Mycobacterium tuber-culosus составляет от 0,0012 до 0,004 мм. Некоторые же бактерии находятся у границы или за пределами разрешающей способности оптического микроскопа. Ничтожно малые размеры бактерий ограничивают представление об их морфологическом разнообразии. Вот основные формы бактерий (рис. 23):
Кокки (Cocci)—клетки шаровидные, округлые. Они могут быть одиночными, но чаще собраны по две (диплококки), по четыре (тетра-кокки) или крупными группами в виде цепочек (стрептококки), гроздей (стафилококки), прямоугольных групп (сарцины) и др.
Бациллы (Bacillus) —клетки в виде прямых длинных палочек.
Бактерии (Bacterium)—клетки в виде коротких палочек.
Вибрионы (Vibrio) —клетки слабо закручены в виде запятой.
Спириллы (Spirillum) — клетки спирально закручены в два-три завитка.
Спирохеты (Spirochaetae) —клетки более длинные, чем у спириллы, и закрученные.
Значительную и разнообразную группу составляют подвижные формы бактерий. Органы движения — жгутики в числе от 1 — 2 до многочисленных — располагаются по-разному, что определяет классификацию подвижных форм бактерий на монотрихи, амфитрихи, лофотрихи, перитрихи (рис.23). Жгутики настолько малы, что их видно только в электронном микроскопе или с помощью специального окрашивания — в оптическом.
Размножение и его темпы. У бактерий наблюдается очень простой способ размножения путем дробления клетки или простого деления. Известны немногие группы, размножающиеся путем почкования. Половой процесс и митотическое деление клеток у большинства отсутствуют. Дробление удлиненных форм обычно осуществляется перпендикулярно к их продольной оси. Деление происходит очень быстро, обычно в течение 20— 30 мин (у Высших растений этот процесс протекает 90—120 мин). Расчеты показывают, что одна бактерия (2 мк длины и 1 мк ширины) при делении каждые 36 мин может образовать за 24 ч 636 млн. клеток. Общая длина этих клеток составила бы 33 м. Через два дня они образовали бы массу, объем которой был бы 442 смъ, а общая их длина составляла бы 563 км. На третий день их масса достигла бы 7,5 м3, а вытянутые в длину они могли бы 14 раз опоясать земной шар по экватору. Столь быстрое размножение бактерий ограничивают реальные условия жизни: не соответствующие климатические (погодные) условия, недостаток пищи и, конечно, санитарные мероприятия. Некоторые палочковидные бактерии переживают неблагоприятные условия, образуя одноклеточные эндогенные споры. Споры некоторых видов выдерживают кипячение при 240° С. Однако многие бактерии, в том числе и патогенные (вызывающие болезни), не выносят солнечного света.
Классификация. Классификация бактерий сопряжена со значительными трудностями. Существует несколько подходов к их классификации. В настоящее время отдел Бактерии обычно подразделяют на 4 класса:
1. Класс Бактерии — Bacteria. Объединяет палочковидные неподвижные формы. Многие из них — возбудители опасных болезней: дифтерии, проказы, туберкулеза и др.

Рис. 23. Бактерии — Bacteriophyta. А — кокки;
Б — бациллы; В — спирохеты; Г — различные
формы жгутиковых бактерий:
/ — монотрих, 2 — амфитрих, 3—4 — лофотрих, 5 — перит-рих
II. Класс Миксобактерии — Myxobacteria. Охватывает подвижные палочковидные клетки, лишенные жгутиков. Движение осуществляется путем отталкивания с помощью слизи, выделяемой из конца клетки. Мик- собактерии иногда имеют клеточные ядра.
III. Класс Спирохеты — Spirochetae. Объединяет организмы с мно- гими завитками и заостренными концами. Большинство их имеет про дольную перепонку.
IV. Класс Актиномицеты (лучистые грибы) — Actinomycetes. В нем сосредоточено значительное число видов, у которых как бы соединены признаки бактерий и грибов. В морфологическом отношении актиноми цеты разнообразны, но преимущественно имеют форму тонких, радиаль- но разрастающихся нитей моноподиального ветвления. Сформированных ядер нет. Во многих руководствах актиномицеты относятся к несо- вершенным грибам. Некоторые актиномицеты представ ляют большой интерес для промышленного производства антибиотиков: биомицина, стрептомицина, тетрациклина и др.
Жизнедеятельность бактерий. Большинство видов бактерий — гетеротрофные организмы, получающие необходимые им питательные вещества и энергию при разложении отмерших, растений и животных и потому являющиеся сапрофитами. Меньшая часть существует за счет живых организмов (паразиты), зачастую вызывая различные заболевания человека, животных и растений (патогенные бактерии). Развиваясь на том или ином субстрате, бактерии накапливают в своем теле необходимые запасные вещества в виде гликогена, жиров, белков и др. Часть видов для своего развития нуждается в свободном кислороде (аэробные бактерии), другие развиваются в бескислородной среде (анаэробные бактерии). Лишь небольшую группу представляют автотрофные организмы, содержащие бактериохлорофилл и другие пигменты, а потому способные к фотосинтезу, а также хемотрофные, осуществляющие синтез органических веществ из углекислого газа и воды за счет энергии, освобождающейся при окислении ими различных веществ (см. стр. 138).
Методы исследования бактерий. Для установления вида бактерий недостаточно микроскопически исследовать их форму, т. к. морфологическое разнообразие их невелико. Первостепенное значение имеет изучение физиологических особенностей бактерий, их способности неодинаково воздействовать и реагировать на разные вещества. Для этого используют искусственные культуры бактерий на разных питательных средах. Такими средами служат мясопептонный бульон, желатина, агар-агар с добавлением к ним других питательных веществ и иные среды. Желатина, агар-агар при остывании образуют студни. Питательную среду в пробирках, закупоренных ватой, или в чашках Петри стерилизуют в автоклаве. Тот или иной материал, содержащий бактерии, особой прокаленной иглой вносят на питательную среду после ее остывания. Пробирки или чашки Петри с посевом устанавливают в термостат — прибор в виде шкафа, где поддерживается постоянная температура, благоприятная для размножения бактерий. Последние, размножаясь, образуют колонии, форма которых, окраска, способность разжижать твердую питательную среду и другие признаки специфичны для определенных видов. Бактерии, взятые тем же способом из одной колонии и перенесенные в новые сосуды, могут образовывать колонии только одного вида. Такие колонии называют чистыми культурами. Культуры бактерий исследуют под микроскопом, применяя различные способы окраски и другие приемы микроскопической техники. Чистые культуры бактерий соответствующих видов имеют широкое применение при сквашивании молока, при силосовании и в целом ряде других технологических процессов.
Значение бактерий. Бактерии распространены повсеместно на всей планете. Они встречаются на поверхности почвы и в почве, в глубоких
слоях земной коры, в воде, в теле мертвых и живых растений и животных и даже в атмосфере. Конечно, концентрация бактерий на Земле неравномерна и зависит от разных условий. Так, зимой в городах в 1 м3 воздуха насчитывается около 4,5 тыс. бактерий, а летом — от 10 до 25 тыс. В 1 г бедной органическими веществами почвы содержится до 500 млн. бактерий, тогда как в богатой их количество достигает 6 млрд. В природе и жизни человека бактерии играют важную роль: иногда положительную, а иногда и резко отрицательную.
Гниение — процесс разложения белковых соединений, приводящий к минерализации остатков растений и трупов животных. Он состоит из нескольких последовательных фаз, причем каждая из них обычно совершается под воздействием определенной группы бактерий. Благодаря гниению (и брожению) сложные органические вещества превращаются в простые, в результате чего осуществляется «круговорот веществ» в природе. Минерализованные вещества становятся вновь доступными для высших растений и могут поглощаться корнями последних.
Особое значение для зеленых растений имеют процессы аммонификации и нитрификации, происходящие в почве, навозе под воздействием бактерий — аэробных хемосинтетиков, обогащающих почву связанными, доступными для растений формами азота (см. стр. 142).
Связывание свободного атмосферного азота. Этот процесс достигает в природе значительных масштабов и осуществляется азотфиксирующими бактериями-аэробами и в меньшей степени анаэробами.
Важная роль принадлежит клубеньковым бактериям. Эти почвенные бактерии, как уже отмечалось, внедряются в корни бобовых, где за счет атмосферного азота создают белковые компоненты своего тела. После отмирания бактерий эти вещества усваивает растение-хозяин. В пожнивных остатках бобовых содержится много азотистых веществ, поэтому бобовые служат хорошими предшественниками других сельскохозяйственных культур. Подсчитано, что на 1 га посева бобовых в среднем связывается около 200 кг азота.
Брожение — процесс разложения бактериями безазотистых веществ (углеводов и др.).
Брожение молочнокислое: конечный продукт — молочная кислота (СзНбО3), получаемая в результате деятельности молочнокислых бактерий — Lactobacterium acidophilum и др. Накопление в субстрате, подвергающемся брожению, молочной кислоты, препятствует развитию гнилостных и других бактерий. Молочнокислое брожение широко используется в практике при сквашивании молока, силосовании кормов и других процессах.
Брожение маслянокислое: конечный продукт — масляная кислота (С4Н8О2), получаемая в результате деятельности маслянокислых бактерий — Clostridium pasteurianum и др. Широко распространено в почве при разложении растительных остатков. Маслянокислое брожение используется при приготовлении сыров, им обусловлено также прогоркание масла.
Брожение уксуснокислое: конечный продукт — уксусная кислота (С2Н4О2), получаемая в результате деятельности уксуснокислых бактерий — Acedobacter aceti и др. Применяется для получения уксуса из вина.
Большую роль играют также пектинобактерии, например Clostridium pectinovorum и другие, вызывающие пектиновое брожение. Они разлагают пектиновое межклеточное вещество, вызывая мацерацию клеток, что имеет большое значение в процессе мочки льна, как способствующее отделению волокна от других тканей. В почве присутствуют также различные бактерии, сбраживающие клетчатку и участвующие в процессе почвообразования.
Особая роль в природе принадлежит хемосинтезирующим серобактериям и железобактериям. Серобактерии используют сероводород, вы-ляющийся при гниении, окисляя его до серы, а последнюю — до серной слоты, которая образует сернокислые соли, доступные для зеленых г астений. Железобактерии добывают необходимую для синтеза энергию путем окисления закисных форм железа в окисные. Железобактерии многочисленны в некоторых источниках, болотах, где часто служат присной образования залежей болотной железной руды.
Вообще в жизни почвы как природноисторического тела важная роль принадлежит почвенным бактериям. Необходимо еще раз отметить особое, чисто сельскохозяйственное значение бактерий в процессе образования почв и повышения их плодородия. Некоторые бактерия используются в качестве удобрения (азотобактерин, фосфобактерин, нитрагин и др.).
В последние годы бактерии, особенно актиномицеты., приобретают большое значение как продуценты многих антибиотиков, все более широко используемых в медицине, ветеринарии и в мероприятиях по защите растений от болезней и вредителей.
Патогенные бактерии, проникая тем или иным путем в тело человека, животных и растений, могут вызывать тяжелые заболевания, выделяя различные ядовитые вещества — токсины. Некоторые бактерии отличаются очень высокой токсичностью. В этом отношении наиболее показателен выделяемый некоторыми бациллами, например Clostridium botulinum, нитротоксин ботулизма. Лучшая среда для активной жизнедеятельности этих бацилл и накопления токсического вещества — колбаса (botulum), сырое мясо, рыба и даже овощные консервы. Токсины ботулизма — это белки, молекулы которых слабо расщепляются ферментами пищеварительной системы. Токсичность нитротоксина ботулизма просто фантастична: 1 г вещества может убить 60 млрд. мышей (1 200 000 т живого веса!).
Широко известны такие болезни человека, как ангина, брюшной тиф, туберкулез, холера и многие другие, а у животных — сибирская язва, паратиф, актиномикоз, сап и другие, вызываемые соответствующими патогенными бактериями. Многие из этих заболеваний — общие для человека и животных.
От момента внедрения патогенных бактерий в организм до проявления болезни в большинстве случаев проходит некоторый период (инкубационный), во время которого бактерии размножаются. Организм вырабатывает особые вещества — антитела и антитоксины, обезвреживающие бактерии и их токсины. При благоприятном исходе болезни организм может приобретать иммунитет по отношению к повторной инфекции бактериями данного вида. На этом основано применение лечебных и предупредительных вакцин, получаемых из убитых или ослабленных бактерий, а также лечебных сывороток, содержащих эти антитела.
У высших растений также наблюдаются заболевания, вызванные бактериями — бактериозы. Однако растения менее подвержены им в силу защищенности клеток плотными стенками и кислой реакции клеточного содержимого, неблагоприятной для развития бактерий. Бактериозы поражают многие культурные растения. Так, у картофеля мокрая и сухая гниль клубней вызывается Bacillus solaniperda и др. Бактериозами поражаются капуста (Bacterium campestre), свекла (Bacterium betae), томаты (Phytobacter lycopersicum). Бактериальный рак у яблони, груши, персика и других вызывает Bacterium tumefaciens. Возбудители проникают в тело растений при различных повреждениях тканей в первую очередь всевозможными вредителями: насекомыми, червями и др. Меры борьбы с бактериозами, как и с другими болезнями растений, составляют предмет фитопатологии. Большое значение имеет выведение сортов, устойчивых к бактериозам
Средства борьбы с бактериями. Патогенные, гнилостные и другие бактерии угрожают здоровью человека и животных или вызывают порчу пищевых продуктов, кормов, различных материалов. Меры борьбы : неблагоприятной микрофлорой следующие:
Дезинфекция — воздействие различными ядовитыми для бактерий веществами: сулемой, карболовой кислотой, формалином, хлорной известью, спиртом и др.
Стерилизация — уничтожение бактерий высокими температурами. Обычное кипячение при 100°С не гарантирует полного уничтожения микроорганизмов, особенно их спор. Лишь нагревание до 120—130°С под высоким давлением в специальном приборе — автоклаве в течение 20— 30 мин убивает не только бактерии, но и их споры. Кипячение, повторяемое ежедневно, в течение нескольких дней, может привести к полной стерилизации. Прокаливание инструментов, посуды также достигает этой цели.
Пастеризация — нагревание до температуры 65—95°С, которое приводит к гибели большинства гнилостных, патогенных и других бактерий, ко не спор и широко применяется для недлительного сохранения молока и других продуктов.
 Отрицательное
значение многих бактерий, особенно
патогенных, настолько велико, что
давно уже возникла необходимость в
государственных и даже международных
мероприятиях по защите здоровья людей,
растений и животных.
Отрицательное
значение многих бактерий, особенно
патогенных, настолько велико, что
давно уже возникла необходимость в
государственных и даже международных
мероприятиях по защите здоровья людей,
растений и животных.
ОТДЕЛ ГРИБЫ — MYCOPHYTA, FUNGI
Насчитывают свыше 100 тыс. видов грибов. Эта группа представляет крупнейший ряд эволюции. Отрасль науки, изучающая грибы, именуется микологией.
По своей морфофизиологической организации грибы обособлены от остального мира живых существ. Происхождение еще не выяснено. Одни микологи считают, что разные группы грибов произошли от разных групп водорослей. Другие считают более вероятным происхождение грибов от каких-то сапрофитных жгутиковых организмов. Родству грибов с водорослями противоречит полное отсутствие у первых пластид, присутствие в клетках гликогена, расположение жгутиков и др. В отличие от водорослей эволюция грибов шла по пути приспособления к гетеротрофности — сапрофитизму и паразитизму.
Таблица 1. Систематика и основные признаки грибов
Ц арство
Fungi
арство
Fungi
О бщая
характеристика
бщая
характеристика
Питание гетеротрофное, так как у них нет хлорофилла, и поэтому они не способны к фотосинтезу. Грибы могут быть: паразитами, сапрофитами или симбионтами. Жесткая клеточная стенка содержит хитин, формирующий фибриллы; исключение составляют Oomycota, у котрых обнаружена целлюлоза.
Тело обычно представлено мицелием, т.е. состоит из сети трубчатых нитей, которые называются гифами. Если запасают углеводы, то, как правило, в виде гликогена, а не в виде крахмала. Размножаются спорами. Неподвижны.
Отдел Oomycota |
Отдел Zygomycota |
Отдел Ascomycota |
Отдел Basidiomicota |
Половое размножение путем оогамии, когда оосфера (женская гамета) сливается с мужской гаметой, и образуется ооспора
|
Половое размножение путем конъюгации, когда два гаметангия сливаются с образованием зигоспоры
|
При половом размножении внутри специальной структуры, которая называется сумкой или аском, образуются споры (аскоспоры)
|
При половом размножении образуются базидии, к которым снаружи прикреплены споры (базидиоспоры)
|
Бесполое размножение посредством зооспор, образующихся в спорангиях |
Бесполое размножение при помощи конидий или спорангиев, содержащих споры. Зооспор нет. |
Бесполое размножение конидиями. Спорангиев не образуют |
Бесполое размножение спорами. Встречается не часто |
Гифы нечленистые |
Гифы нечленистые; мицелий обширный, хорошо развитый, ветвящийся |
Гифы членистые |
Гифы членистые |
Примеры: Phytophthora infestans -факультативный паразит, возбудитель картофельной гнили; Pythium - многие опасные факультативные паразиты; некоторые вызывают выпревание (черную ножку) проростков; Peronospora - облигатные паразиты, вызывающие ложную мучнистую росу у крестоцветных, например у капусты |
Примеры: Rhizopus stolonifer-обыкновенная хлебная плесень, сапрофит; Мисог - обычные плесени, сапрофиты |
Примеры: Penicillium и Aspergillus-сапрофитные плесени; Saccharomyces (дрожжи) -одноклеточные сапрофиты; Erysiphe - облигатные паразиты, вызывающие настоящую, мучнистую росу, например у ячменя; Ceratocystis ulmi -паразит, вызывающий голландскую болезнь вяза |
Примеры: Agaricus campestris-шампиньон обыкновенный-сапрофит |
Отдел Chytridiomycota
Небольшая группа микроскопических (чаще всего одноклеточных) грибов. Пример: Synchytrium endobioticum-паразит, вызывающий рак картофеля
\У Отдел Deuteromycota (Fungi Imperfecti)
Грибы, у которых никогда не наблюдается половое размножение и систематика которых до сих пор не выяснена. Пример: Trichophyton, вызывающий грибковые заболевания ног и стригущий лишай.
О братите внимание, что названия отделов имеют окончание -mycota
Особенности морфофизиологической организации. Вегетативное тело грибов называют мицелием, или грибницей. Мицелий состоит из тонких ветвящихся нитей — гиф. У низших грибов гифы лишены перегородок и представляют как бы одну сильно разветвленную гигантскую клетку. У высших грибов они разделены поперечными перегородками на клетки членики). Гифы покрыты стенкой (оболочкой), которая только у немногих низших грибов (оомицетов) состоит из целлюлозы. У большинства же грибов стенка гиф имеет более сложный состав: у низших она состоит из пектиновых веществ, а у высших — из углеводов, близких целлюлозе, и азотистых веществ, сходных с хитином насекомых. Гифы нарастают верхушками. При благоприятных условиях они отличаются очень быстрым темпом роста. У высокоорганизованных грибов гифы редко плотно сплетаются и образуют плектенхиму — ложную ткань, состоит из переплетения нитей, нарастающих верхушками, тогда как настоящая ткань растений состоит из клеток, которые делятся по всем правлениям. Из плектенхимы формируются так называемые «плодовые тела», ризоморфы (шнуры в несколько миллиметров толщиной и длиной, достигающей иногда нескольких метров) и другие образования. У грибов нет приспособлений для проведения воды и защиты от испарения, поэтому они живут во влажных местообитаниях.
Важнейшая черта организации грибов — отсутствие пластид. Ядра очень мелкие, в числе одного -двух или многочисленны. Запасные вещества откладываются в виде гликогена или жира, крахмал никогда не образуется.
Мицелий обычно погружен в субстрат, откуда он поглощает пищу. Большинство грибов питается остатками отмерших растений — сапрофитно. Грибы-сапрофиты образуют ферменты, разрушающие целлюлозные клеточные стенки и лигнин. Значительно меньше сапрофитов питается остатками животного происхождения. Среди грибов-паразитов 10 тыс. видов паразитируют на растениях, менее 1 тыс. — на животных, человеке. Многие грибы часть жизни проводят как паразиты, часть — сапрофиты. Нередко грибы вступают в сожительство с водорослями и даже высшими растениями. В процессе симбиоза могут возникнуть новые образования, например микориза (грибокорень). Симбиоз грибов и водорослей привел к образованию лишайников.
Размножение. В процессе размножения у грибов наблюдаются специфические особенности. У них хорошо выражена редукция полового процесса. В связи с этим выработались весьма разнообразные способы вегетативного и бесполого размножения.
Вегетативное размножение происходит при помощи: частей мицелия - обособленных члеников гиф с тонкими стенками или хламидоспор — клеток с толстыми стенками, на которые распадаются гифы мицелия; почкования, т. е. образования на клетках округлых выростов, отделяющихся от них.
Бесполое размножение осуществляется при помощи различных спор, развивающихся на мицелии: спорангиоспор (образуются внутри спорангиев); зооспор (образуются внутри зооспорангиев); конидиоспор — экзогенных спор, которые формируются по одной или цепочками на конидиях (особых гифах, расположенных вертикально) путем почкования. У некоторых грибов в жизненном цикле несколько форм бесполого спороношения.
Половое размножение. У низших грибов формы полового процесса исключительно разнообразны: хологамия, зигогамия, гетерогамия, изогамия, оогамия. Зигота всегда некоторое время находится в покое, затем прорастает и образует зооспоры или короткую гифу с зооспорангием, спорангием, конидией. Перед прорастанием зигота делится редукционно, следовательно, жизнь почти всех низших грибов проходит в гаплоидной фазе (п хромосом), диплоидна лишь зигота (2п хромосом). У высших грибов формы полового процесса своеобразны. У них ядра противоположных полов почти всегда сливаются не сразу, а сначала располагаются парами, образуя дикарионы, которые делятся синхронно, так что число их увеличивается. После слияния ядер образуется диплоидное ядро, которое делится редукционно, и гаплоидные ядра становятся ядрами спор полового размножения. Следовательно, у высших грибов в жизненном цикле чере-
Рис.
24. Половое спороношение высших
грибов. А — схема образования сумки;
Б — схема образования базидии; 1 —
споры

двух типов (рис. 24)а) аскоспоры -образуются эндогенно, большей частью по восемь в особых клетках, называемых асками, или сумками, где происходит слияние ядер дикариона, а затем редукцинное деление; б) базидиоспоры — образуются экзогенно, большей частью по четыре на поверхности особых клеток, именуемых базидиями. У высших грибов половое спороношение завершает жизненный цикл.
органические вещества, в основном остатки отмерших растений,и делают их доступными
для питания других растений. Грибы, вызывающие спиртовое брожение, широко используются в хлебопечении, для производства вина, спирта, пива, кваса, кефира. Кроме того, дрожжи используют как пищевой, кормовой и лечебный продукты. Шляпочные грибы нередко имеют пищевое значение. Однако стенки гиф плохо перевариваются, поэтому белки грибов плохо усваиваются. Некоторые виды грибов используют для получения антибиотиков. Многие грибы вызывают болезни сельскохозяйственных растений.
Громадный ущерб народному хозяйству наносят грибы, разрушающие древесину. Некоторые виды вызывают болезни у человека и животных, а также отравления. Грибы паразитируют в теле организмов, имеющих кислую реакцию, которую не выдерживает большинство патогенных бактерий.
Классификация. Грибы делят на 4 класса: Фикомицеты, Аскомицеты, Базидиомицеты, Несовершенные грибы. В первом классе объединяют низшие грибы. Они характеризуются отсутствием мицелия или мицелием, не разделенным перегородками на членики. Остальные три класса объединяют в группу высших грибов: их мицелий состоит из членистых гиф.
КЛАСС ФИКОМИЦЕТЫ - PHYCOMYCETES
Общее число видов около 1 тыс. Важнейший признак организации —-отсутствие мицелия или же мицелий, состоящий из нечленистых гиф. В классе Фикомицеты различают три подкласса: Архимицеты, Оомицеты и Зигомицеты.
Подкласс Архимицеты — Archimycetidae
Число видов 300. Вегетативное тело представлено либо в виде голо цитоплазматической массы, которая развивается внутри клеток растения хозяина, либо в виде клетки, от которой отходят слаборазвитые гифы, разделенные перегородками. Бесполое размножение осуществляете: зооспорами, в большинстве случаев снабженными одним жгутиком. Формы полового процесса исключительно разнообразны (хологамия, изогамия, гетерогамия, зигогамия и др.). Живут архимицеты преимущественно в водной среде. Многие паразитируют на водорослях, на других видах грибов, некоторые виды — на высших наземных растениях.
Рассаду капусты в парниках нередко поражает ольпидий — Olpidiun brassicae (рис. 25). Этот паразит вызывает заболевание, известное noд названием «черная ножка». Вегетативное тело гриба — плазмодий (голый комочек цитоплазмы) развивается в клетках эпидермы или паренхимы коры молодых растений. Ядра размножаются делением, затем плазмодий покрывается стенкой и превращается в зооспорангий. На зооспорангии образуется длинная выводная шейка, которая достигает поверхности органа. Через эту шейку зооспоры выходят наружу и попадают в почвенную воду. Одножгутиковые зооспоры приближаются к новому растению, прикрепляются к эпидерме и переливают в ее клетки содержимое. Начинается развитие нового зооспорангия. При благоприятных условиях жизненный цикл длится 2—3 дня. Половой процесс изогамный. Гаметы, сливаясь, образуют планозиготу — зиготу, которая передвигается с помощью жгутиков. Она имеет два ядра. Планозигота одевается толстой оболочкой и зимует в теле растения-хозяина. При прорастании планозиготы ядра сливаются, а затем делятся редукционно, образуется плазмодий, из которого развивается зооспорангий с зооспорами. Меры борьбы — умеренно влажное содержание парников и дезинфекция почвы, например формалином.
Другой важнейший представитель подкласса — возбудитель рака картофеля — Synchytrium endobioticum. Зооспоры паразита, проникая в клетки клубня, вызывают их сильное увеличение. Клетки, окружающие пораженные участки ткани, многократно делятся, что приводит к образованию на клубнях бугристых наростов (рис. 26). Пораженные клубни недоразвиты и содержат мало крахмала. При бесполом размножении в

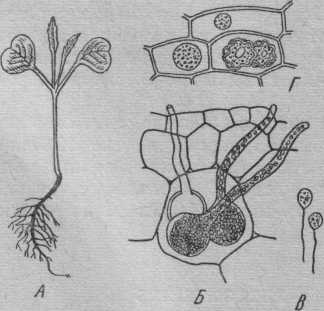
Рис.26.
Синхитрий — Synchytrium
endobioticum.
А — клубни и столоны картофеля,
пораженные раком; Б — цистысях
бугристых наростов (рис.
180
отличие от ольпидия формируется не один, а целая группа зооспорангиев. Половой процесс изогамный. Зигота, снабженная двумя жгутиками, проникает в клубень картофеля, покрывается толстой оболочкой, превращается в цисту и зимует. Циста сохраняет жизнеспособность в течение 10 лет. Рак картофеля широко распространен в США и Западной Европе. Возбудитель этой болезни занесен в СССР во время второй мировой войны. Меры борьбы — обработка почвы соответствующими химикатами, но наиболее перспективна селекция ракоустойчивых сортов картофеля.
Подкласс Оомицеты — Oomycetidae
Представители этого подкласса, как и следующего, имеют хорошо развитый мицелий. Сильно разветвленные гифы лишены перегородок и содержат много ядер. Поперечные перегородки возникают только при отчленении органов размножения. Большая часть жизни оомицетов проходит в гаплоидной фазе. Диплоидна лишь зигота, при прорастании которой осуществляется редукционное деление. Жизненный цикл большинства видов в значительной мере связан с водной средой. Основные признаки оомицетов: 1) оогамный половой процесс, 2) наличие целлюлозы в стенке гиф и отсутствие хитина, 3) образование зооспор; лишь у представителей высшей организации бесполое размножение с помощью конидиоспор.
Фитофтора — Phytophthora infestans (рис.27)—паразит, вызывающий заболевание ботвы картофеля и клубней. Мицелий развивается в мезофилле листьев, что приводит к отмиранию клеток хлорофиллоносной паренхимы. В результате на листьях образуются бурые пятна. Развитие паразита ведет к резкому сокращению зеленой поверхности растения, к сокращению урожая клубней и содержания крахмала в них. Размножается фитофтора, как правило, бесполым путем. Гифы паразита через устьица высовываются наружу, ветвятся и отчленяют овальные конидии,

Рис. 27. Фитофтора — Phytophthora infestans. A — побег картофеля, пораженный фитофторой; Б — разрез пораженного листа, на котором видны гифы гриба и конидии на конидиеносцах, прорастающих через устьица наружу; В, Г — зооспорангий и выход зооспор; Д — зооспоры; Е — прорастание зооспоры; Ж—конидиоспора; 3 — прорастание конидиоспоры; И, К — клубни, пораженные фитофторой; Л, М — мицелий фитофторы в ткани клубня
которые переносятся воздушными течениями и попадают с капельками дождя или росы на листья. Конидии развиваются, как зооспорангии, продуцирующие двужгутиковые зооспоры. Через эпидерму зооспоры проникают внутрь листа и прорастают в мицелий. При недостаточной влажности и высокой температуре конидия прорастает непосредственно в гифу. Заболевание более широко распространяется в дождливую погоду, так как темп размножения зооспорами значительно выше, чем конидиоспорами, Конидии или зооспоры, падая на почву, могут заражать клубни картофеля. Половое размножение наблюдается лишь вне растения-хозяина в почве. На гифах образуются оогонии и антеридии. Оогоний имеет шаровидную форму, внутри него формируется одноядерная яйцеклетка. Гифа с антеридием растет в сторону оогония и стыкуется с ним. Затем выросты антеридия через поры врастают внутрь оогония, достигают яйцеклетки и часть содержимого с одним ядром сливается с яйцеклеткой. Оплодотворенная яйцеклетка покрывается стенкой и превращается в ооспору. По окончании периода покоя ооспора прорастает в гифу с зооспорангием. Ооспоры перед прорастанием делятся редукционно. Зимуют ооспоры, а также мицелий на растительных остатках. Заражение происходит через почву и клубни картофеля. Ph. infestans занесена в Европу из Америки в первой трети XIX в. Меры борьбы: агротехнические мероприятия, опрыскивание бордоской жидкостью, выведение устойчивых сортов.
Многие представители подкласса Оомицеты поражают сельскохозяйственные растения: Plasmopara vitlcola вызывает болезнь винограда — мильдью, P. brassicae — болезнь капусты, Peronospora tabacina — болезнь табака и др.
Подкласс Зигомицеты — Zigomycetidae
Важнейшие признаки: 1) зигогамный половой процесс, 2) наличие хитина и отсутствие целлюлозы в стенках гиф, 3) образование спорангиоспор, конидиоспор и полное отсутствие зооспор, 4) почти все виды ведут наземный образ жизни.
Из зигомицетов общеизвестен род мукор — Мисог. В СССР широко распространен М. mucedo. Этот сапрофит развивается в виде плесени в почве, особенно часто на конском навозе, а также на пищевых продуктах. Размножается преимущественно бесполым способом. На мицелии образуются многочисленные спорангиеносцы со спорангиями (рис.28). Содержимое спорангия распадается на множество многоядерных спор. После созревания оболочка спорангия лопается, и споры рассеиваются. Половой процесс наблюдается сравнительно редко. Он возможен, если рядом окажутся два гетероталличных мицелия (гомоталличные зигомицеты встречаются относительно редко). Гифы мицелиев растут навстречу друг другу, и, наконец, смыкаются. На расширенных концах отделяются многоядерные участки. Стенки гиф в месте их соприкосновения растворяются, и содержимое обеих гиф, в том числе и ядра, сливается. Образовавшаяся зигота покрывается толстой стенкой. После периода покоя зигота прорастает в короткую гифу, увенчанную спорангием. При образовании спор диплоидные ядра делятся редукционно. В неблагоприятных условиях мицелий мукора расчленяется на членики, которые покрываются толстой стенкой и превращаются в хламидоспоры. С наступлением благоприятных условий хламидоспоры прорастают в мицелий. В других случаях мицелий распадается на оидии, которые при условии обильной пищи делятся путем почкования. Оидии в сахарном растворе вызывают спиртовое брожение (мукоровые дрожжи).
10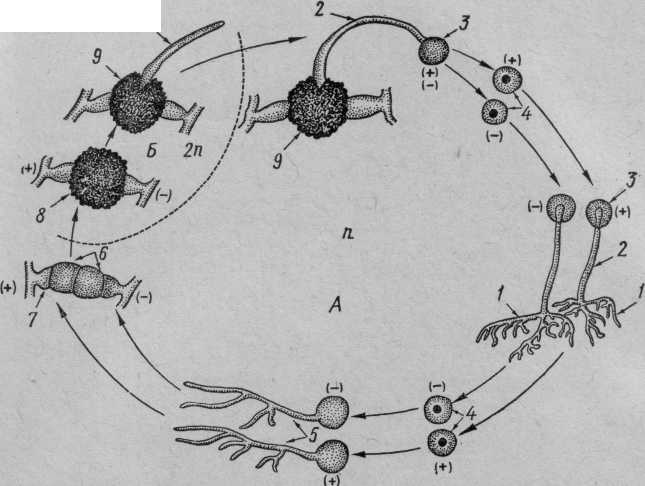
Рис. 28. Жизненный цикл мукора — Mucor mucedo. А — гаплоид; Б — диплоид:
1 — два гетероталличных мицелия, 2 — спорангиеносец, 3 — спорангий, 4 — споры, 5 — прорастание спор, 6 — отделившиеся участки гиф, 7 — подвесок, 8 — зигота, 9 — прорастающая зигота, 10 —
промицелий
КЛАСС АСКОМИЦЕТЫ, ИЛИ СУМЧАТЫЕ ГРИБЫ — ASCOMYCETES
Объединяет около 25 тыс. видов. Аскомицеты имеют развитый мицелий, который состоит из членистых гиф, погруженных в субстрат. На поверхности образуются лишь органы размножения. Большинство видов — сапрофиты. Они живут в почве, питаясь остатками отмерших растений, на пищевых продуктах. Однако известны и паразиты растений, реже животных и даже человека. Некоторые виды начинают жизненный цикл как паразиты, а заканчивают как сапрофиты.
Бесполое размножение осуществляется конидиоспорами. Половой процесс у примитивных представителей заключается в слиянии двух члеников мицелия. Зигота не проходит периода покоя, а диплоидное ядро сразу же делится редукционно, а затем еще раз митотическ|1. В результате образуются 8 гаплоидных аскоспор. У большинства высокоорганизованных аскомицетов образуются органы полового размножения: женский — архикарп и мужской — антеридий. Архикарп состоит из двух клеток: нижней вздутой — аскогона и верхней цилиндрической — трихогины (рис. 29). Антеридий состоит из одной цилиндрической клетки. Оба органа многоядерны. Антеридий смыкается с верхушкой трихогины, содержимое его переливается в архикарп, и происходит слияние цитоплазм. Ядра при этом не сливаются, но сближаются попарно, образуя дикарионы. Дикарион состоит из двух ядер разного происхождения: аскогонного (женского) и антеридиального (мужского). После формирования дикарионов из аскогона начинают развиваться ветвящиеся аскогенные гифы. В них переходят дикарионы, которые затем делятся, образуя новые, дочерние дикарионы. Соответственно образованию новых дикарионоз в аскогенных гифах возникают перегородки, каждый членик содержит по одному дикариону. В завершающей фазе полового процесса в верхушечных члениках аскогенных гиф происходит слияние ядер дикарионов
.

Рис. 29. Половой процесс у аскомицетов — Ascomycetes. Л — мицелий; Б — органы полового размножения; В — срастание трихогины и антеридия и переход протопласта последнего в аскогон; Г — образование в аскогоне дикарионов; Д — развитие аскогенных гиф; Е — образование сумки: 1 — архикарп, 2 — аскогон, 3 — трихогина, 4 — антеридий, 5 — аскогенные гифы, 6 — дикарион на верхушке аскогенной гифы, 7—12 —схема завершения полового процесса, 13 — образование зиготы,14—15 — редукционное деление и последующее развитие сумки с аскоспорами
Зигота тут же редукционно делится, в результате чего получается 8 (иногда 4) гаплоидных клеток — аскоспор. Верхушечные членики, в которых содержатся аскоспоры, разрастаются, их называют сумками, или асками. Следовательно, у высокоорганизованных аскомицетов чередуются три фазы: гаплоидная (основная), дикарионная и очень непродолжительная диплоидная. Между асками всегда находятся «стерильные» гифы. Они называются парафизами, Парафизы выполняют буферную роль, но, возможно, способствуют рассеиванию аскоспор. Аски и парафизы образует гимениальный слой, или гимений. Гимениальный слой аскомицетов расположен на плотном сплетении гиф, именуемом плодовым телом. В большинстве случаев плодовое тело более или менее окружает гимений. У более просто организованных представителей плодовые тела не развиваются, сумки возникают прямо на гифах мицелия. В зависимости от наличия или отсутствия плодового тела класс Аскомицеты подразделяют на два подкласса: Голосумчатые и Плодосумчатые.
Подкласс Голосумчатые — Protoascomycetidae, или Gymnoaseomycetidae
Характеризуются отсутствием плодовых тел и гимениальиого слоя. Аски образуются непосредственно на мицелии или в отдельных неспециализированных клетках.

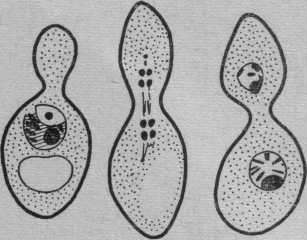
Рис. 30. Дрожжи, род Saccharomyces. А — клетка пивных дрожжей — 5. cerevisiae Б — то же в стадии почкования; В — последовательные стадии деления винных дрожжей — S. ellipsoideus: 1 — ядро, 2 — ядрышко, 3 — вакуоли, 4 — митохондрии, 5 — гликоген
Наиболее широко распространены виды рода дрожжи — Saccharomyces, например пивные, или хлебопекарные, дрожжи — 5. cerevisiae. Они вызывают спиртовое брожение и имеют большое практическое значение в пивоваренной промышленности, хлебопечении, производстве спирта. Известны только в культуре. Тело состоит из одиночных овальных клеток с одним ядром. Вегетативное размножение осуществляется почкованием (рис.30). В неблагоприятных условиях происходит половой процесс, в результате которого внутри клетки развивается сумка с 4 или 8 аскоспорами. В виноделии используют другой вид — винные дрожжи — S. vini или S. ellipsoideus, который живет на поверхности или в мякоти сочных плодов
.

Рис. 31. Плодовые тела аскомицетов — Ascomycetes. А—Б — клейстогеций; В — Г — перитеций; Д—Е — апотеций (А, В, Д — внешний вид, Б, Г, Е — в разрезе, сильно увеличено)
кислоты. Бесполое размножение — посредством конидиоспор. Конидиеносец не разделен на членики, на вершине шаровидно вздут.
Некоторые представители' из рода пеницилл — Penicillium, например P. notation, P- chrysogenum и другие, служат для производства антибиотика пенициллина. Виды P. roqueforti и P. camemberti используют

в производстве некоторых сортов сыра. Бесполое размножение так же посредством конидиоспор, расположенных в виде кисточки на верхнем конце членистого конидиеносца (см. рис.32). Половое размножение бывает редко.
Рис. 32. Пеницилл —род Penicillium (А); аспергилл род Aspergillus (Б):
/ — членистый конидиеносец с конидиоспорами, 2 — одноклеточный конидиеносец с конидиоспорами, 3 — разрез клейстотеция
КЛАСС БАЗИДИОМИЦЕТЫ — BASID1OMYCETES
 Общее
число видов около 30 тыс. Основные
признаки: мицелий состоит из членистых
гиф, специальных органов полового
размножения нет, половой процесс
осуществляется путем слияния члеников
двух гаплоидных гиф. У гетероталличных
базидиомицетов, которых большинство,
сливаются гифы мицелиев различных
особей с противоположным знаком (+ и
—). Однако слияние содержимого члеников
не сопровождается слиянием ядер.
Образуются дикарионы, которые размножаются
делением. Половой процесс завершается
слиянием ядер дикарионов, редукционным
делением и образованием базидии с
четырьмя базидиоспорами. Базидия
гомологична сумке аскомицетов (рис.29).
В жизненном цикле базидиомицетов
доминирует дикарионная фаза, а гаплоидная
и диплоидная очень кратковременны.
У большинства видов базидиоспоры
образуются на плодовых телах, форма
и размер которых очень разнообразны.
Гимений чаще всего располагается на
нижней стороне плодового тела. Он
состоит из базидий, псевдопарафиз и
цистид. Псевдопарафиза — членик
гифы с дикарионом. Цистида — более
крупный, чем псевдопарафиза, членик,
возвышающийся над гимениальным слоем.
Форма псевдопарафиз и цистид для многих
видов достаточно постоянна и служит
важным признаком для их определения.
Поверхность плодового тела, несущая
гимекий, называется гименофором. У
примитивных представителей он гладкий,
а у высокоорганизованных имеет форму
зубцов, пластинок, трубочек (рис. 33), что
значительно увеличивает поверхность
гимениального слоя. У некоторых видов
базидии развиваются прямо на мицелии.
Общее
число видов около 30 тыс. Основные
признаки: мицелий состоит из членистых
гиф, специальных органов полового
размножения нет, половой процесс
осуществляется путем слияния члеников
двух гаплоидных гиф. У гетероталличных
базидиомицетов, которых большинство,
сливаются гифы мицелиев различных
особей с противоположным знаком (+ и
—). Однако слияние содержимого члеников
не сопровождается слиянием ядер.
Образуются дикарионы, которые размножаются
делением. Половой процесс завершается
слиянием ядер дикарионов, редукционным
делением и образованием базидии с
четырьмя базидиоспорами. Базидия
гомологична сумке аскомицетов (рис.29).
В жизненном цикле базидиомицетов
доминирует дикарионная фаза, а гаплоидная
и диплоидная очень кратковременны.
У большинства видов базидиоспоры
образуются на плодовых телах, форма
и размер которых очень разнообразны.
Гимений чаще всего располагается на
нижней стороне плодового тела. Он
состоит из базидий, псевдопарафиз и
цистид. Псевдопарафиза — членик
гифы с дикарионом. Цистида — более
крупный, чем псевдопарафиза, членик,
возвышающийся над гимениальным слоем.
Форма псевдопарафиз и цистид для многих
видов достаточно постоянна и служит
важным признаком для их определения.
Поверхность плодового тела, несущая
гимекий, называется гименофором. У
примитивных представителей он гладкий,
а у высокоорганизованных имеет форму
зубцов, пластинок, трубочек (рис. 33), что
значительно увеличивает поверхность
гимениального слоя. У некоторых видов
базидии развиваются прямо на мицелии.
Рис. 33. Ложный трутовик — Forties igniarius
( А) белый гриб — Boletus edulis (Б): 1 — плодовое тело, 2 — поперечный срез трубчатого гименофора
Базидиальные грибы подразделяют на два подкласса: Холобазидиомицеты и
Фрагмобазидиомицеты.
Группа порядков Гименомицеты — Hymenomycetidae. Плодовые тела различной формы. Гимений лежит на поверхности плодового тела открыто или обнажается до созревания спор. Большинство живет сапрофитно, паразитов мало. Многие виды вступают в симбиоз с корнями древесных пород, образуя микоризу.
Важное значение имеют представители порядка Афиллофоровые — Aphyllophorales. Сюда относят до 20 семейств. Эти виды с гладким, складчатым или трубчатым гименофором, причем гименофор не отделяется от мякоти плодового тела. Плодовые тела часто кожистые или деревянистые, копытовидные, не загнивающие. Важнейшие виды: настоящий трутовик — Forties fomentarius, ложный трутовик — F. igniarius (см. рис.33), лиственничный трутовик, паразитирующий на древесных растениях, —F. laricis. Некоторое медицинское значение имеет паразитирующий на березе гриб чага — Inonotus obliquus. Домовый гриб — Merulius lacrimans питается только целлюлозными стенками клеток древесины. При благоприятных условиях способен очень быстро разрушать деревянные конструкции мостов, жилых домов. Гриб причиняет большой ущерб на складах древесины, вызывая ее разрушение. Мицелий располагается как внутри субстрата, так и на поверхности. Плодовые тела образуются редко, гименофор складчатый. Распространяется спорами и участками мицелия.
Меры борьбы: постройка сооружений из сухой древесины, удаление и сжигание пораженной древесины, пропитывание древесины специальными антисептическими препаратами.
Порядок Агариковые — Agaricales. Объединяет наиболее широко известные семейства грибов — Пластинниковые — Agaricaceae и Болетусовые — Boletaceae. Это преимущественно почвенные сапрофиты. Общее число видов около 11 тыс. К ним относится подавляющее большинство видов съедобных грибов, а также многие микоризообразователи.
Семейство Пластинниковые включает около 10 тыс. видов. Обычно плодовое тело дифференцировано на ножку, или пенек, и шляпку. Гименофор находится на нижней стороне шляпки в виде радиально расположенных пластинок. Представители: шампиньон двуспоровый—Agaricus bisporus (часто культивируют в специальных теплицах), груздь — Lacta-rius resimus, рыжик — L. deliciosus, опенок осенний — Armillariella mel-lea (развивается как сапрофит на отмершей древесине, но иногда паразитирует). Из числа ядовитых представителей важно отметить красный мухомор — Amanita muscaria и бледную поганку — A. phalloides.
У видов семейства Болетусовые гименофор трубчатый (см. рис.33). Обычно он легко отчленяется от мякоти плодового тела — шляпки. Наиболее богат видами род болетус — Boletus. Некоторые из них имеют промышленное значение: белый гриб — В. edulis, подберезовик — В. scader, дубовик — В. luridus, масленок — В. luteus и др.
Группа порядков Гастеромицеты — Gasteromycetidae. Плодовые тела совершенно замкнутые. Внутри находятся полости, выстланные гиме-нием. После созревания базидиоспор покровы плодового тела разрушаются. Почти все представители — сапрофиты. Плодовые тела очень разнообразны по форме, развиваются то в почве, то на ее поверхности. Представитель порядка — дождевик гигантский — Lycoperdon giganteum. Плодовое тело имеет диаметр до 30 см.
Подкласс Фрагмобазидиомицеты — Phragmobasidiomycetidae
Базидии разделены перегородками на четыре членика. Такие базидии называются фрагмобазидиями. В составе подкласса 4 порядка, из которых ниже рассматриваются два: Головневые и Ржавчинные.
Порядок Головневые — Ustilaginales. Плодовые тела не образуются. Все представители — паразиты высших растений и вызывают их заболевания. Многие паразитируют на хлебных злаках, разрушая зерновки, а также соцветия. Последние густо покрываются целым слоем спор и имеют как бы обожженный вид. Отсюда и название — «головневые» грибы.
На пшенице паразитирует твердая, или вонючая, головня — Tilletia caries, на овсе — Ustilago avenae, на ячмене—U. hordei, на просе — Sphacelotheca panici-miliacei (рис.34). Это наиболее злостные паразиты хлебных злаков. Все они имеют сходный жизненный цикл. Во время обмолота споры паразита прилипают к зерновкам и при посеве попадают в почву. Заражение происходит при прорастании зерновки, когда одновременно прорастает и спора. Ядра дикариона, содержащегося в споре, сливаются. Образовавшееся диплоидное ядро сразу же делится редукционно в споре или в развивающейся из нее базидии. Базидиоспоры не способны заражать растения. Две гетероталличные базидиоспоры сливаются и образуют клетки с дикарионами, из которых развивается мицелий. Этот дикарионный мицелий проникает в конус нарастания растения-хозяина и развивается вместе с ним. До выколашивания растения внешне не отличаются от здоровых. Далее мицелий проникает в формирующееся соцветие, быстро развивается, а затем распадается на массу двуядерных члеников — головневых спор, покрытых толстой стенкой. В результате из влагалища появляется сильно деформированное соцветие. Таким образом, полный цикл развития завершается в течение одного вегетационного периода. У твердой головни пшеницы — Т. caries базиди; не разделены на членики, мицелий проникает в соцветие, сильно разрастается внутри зерновок и поглощает их содержимое. Околоплодник и семенная кожура не разрушаются, и зерновки сохраняют внешнюю форму, но внутри заполнены головневыми спорами. Меры борьбы: обеззараживание посевного материала раствором формалина, опыление мышьяковистыми, ртутными или сероорганическими препаратами,
Более сложный жизненный цикл имеют пыльная головня пшеницы — Ustilago tritici, пыльная головня ячменя—U. nuda и др. Споры попадают на рыльце пестика, где прорастают и образуют фрагмобазидию. Однако базидиоспоры не возникают, а происходит попарная копуляция члеников фрагмобазидий, которые затем прорастают в дикарионный мицелий. Последний проникает в завязь, развивается внутри эндосперма : зародыша, но не разрушает их. Пораженные зерновки не отличаются здоровых ни формой, ни удельным весом и способны прорастать. Следовательно, здесь заражение происходит не в почве, а во время цветения формирования зерновки. При посеве, когда зародыш трогается в рост, когда начинается интенсивный рост мицелия. Особенно сильно он развиваетется в зоне соцветия, где накапливается много питательных веществ. Здесь мицелий распадается на отдельные членики, которые покрываются прочной стенкой и превращаются в споры. При этом соцветие деформируется. Особенно сильно поражаются цветки и кроющие чешуйки. Нередко соцветия настолько разрушаются, что представляют собой черную пылящую массу, и сохраняется лишь главная ось соцветия. Споры распространяются ветром и, попадая на рыльца здоровых растений, прорастаю: Таким образом, полный цикл завершается в два вегетационных период. Меры борьбы затруднены, так как мицелий сохраняется внутри зерновки. Применяют термическую обработку посевного материала, агротехнические мероприятия. Наиболее радикальные меры борьбы — выведение сортов, устойчивых к пыльной головне, и получение посевного материала с,участков, где ее не было.

Рис. 192. Пыльная головня овса — Ustilago avenae (А); пыльная головня проса — Sphacelotheca panici-miliacel {Б); пыльная головня пшеницы —Ustilago tritici (В)\ твердая головня пшеницы-— Tilletia caries (Г); пузырчатая головня кукурузы — Ustilago maydis (Д):
1— спора пыльной головни, 2 —прорастание споры пыльной головни, 3 —спора твердой головни, 4 — проросшая спора твердой головни, 5 — зерновка пшеницы, заполненная спорами твердой головни {головневый мешочек), 6 — прорастание споры пузырчатой головни