

Глава V.
ПОБЕГ И СИСТЕМЫ ПОБЕГОВ
ОБЩАЯ ХАРАКТЕРИСТИКА ПОБЕГА
Определение и терминология. Побег, как и корень,— основной орган высшего растения. Вегетативные побеги выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные, обеспечивающие размножение.
В современной морфологии растений побег в целом принимают за единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение: он с самых ранних этапов развития расчленяется на специализированные части. Вегетативный побег состоит из о с и (стебля) и листьев — в типичном случае плоских боковых органов, сидящих на оси. Ни стебель без листьев (хотя бы зачаточных или рудиментарных), ни листья без стебля (хотя бы укороченного до предела) образоваться не могут. Кроме того, обязательной принадлежностью побега являются почки обеспечивающие длительное нарастание побега и его в е т в л е н и е, т. е. образование системы побегов. Главную функцию побега — фотосинтез — осуществляют листья; стебли — преимущественно несущие органы, выполняющие механическую и проводящую, а иногда и запасающую функцию.
Единство побега и взаимосвязь всех его частей превосходно проявляются в случае резких метаморфозов. Например, при образовании клубня у картофеля видоизменяется весь побег: осевая часть его разрастается в толщину, а листья редуцируются, превращаясь в «бровки»; почки (глазки) сохраняют нормальное строение и расположение (рис. 222).
При образовании луковицы стеблевая часть побега (его ось) разрастается только в ширину, образуя плоское или конусовидное «донце», а сидящие на нем листья в подземной части утолщаются, превращаясь в мясистые, сочные луковичные чешуи; почки занимают соответствующие позиции на морфологической верхушке (в центре) донца и в пазухах чешуевидных листьев (рис. 225). Совершенно ясно, что называть клубни и луковицы видоизменениями стебля, как это иногда делают, неверно: все это — видоизменения сложного органа — побега.
Главная черта, отличающая побег от корня,— его облиственность, а следовательно, наличие узлов. Узлом принято называть участок стебля на уровне отхождения листа или мутовки листьев. У некоторых групп растений (например, злаки, гвоздичные, хвощи) узлы резко обозначены в виде утолщений на стебле; у других растений границы узлов более или менее условны. Если лист или мутовка листьев полностью окружают стебель основанием, то узел называют закрытым, в отличие от открытого узла, который несет лист, не охватывающий его целиком (рис. 115, /, 2, 3).
Участки стебля между соседними узлами называют междоузлиями. •
Обычно на побеге имеется несколько, иногда много узлов и междоузлий, они повторяются вдоль оси побега.

Рис. 115. Метамерность побегов:
/—закрытый узел побега крапивы с двучленной мутовкой листьев — супротивное листорасположение; 2— закрытый узел побега традесканции, полностью охваченный влагалищем листа; 3— открытый (неполный) узел побега липы; 4 - общая схема развития метамеров побега; Пр — прилистники.
Первый побег растения — его главный побег, или побег первого порядка. Он образуется из зародышевого побега, представленного гипокотилем, семядолями, отходящими от семядольного узла, и почечкой. Из неё в дальнейшем формируются все последующие метамеры главного побега. По положению эта почка — верхушечная; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров.
Кроме верхушечной, на побеге образуются боковые почки. У семенных растений они, как правило, находятся непосредственно над узлами, ложен ряд вегетативных метамеров, а конус нарастания превращен в зачаточный цветок или соцветие (вегетативно-генеративные почки иногда называют смешанными, что неудачно). Такие почки обычны для травянистых растений (например, копытень, рис. 117, 4), но бывают и у древесных (например, сирень, бузина, рис. 117, 2). Чисто генеративные, или цветочные, почки заключают в себе только зачаток соцветия, без зеленых ассимилирующих листьев (например, вишня, рис. 117, 5), или одиночный цветок; в последнем случае почку называют бутоном.
Довольно часто наружные листья почки представляют собой специализированные почечные чешуи, выполняющие защитную функцию и предохраняющие части почки от высыхания. Иногда пишут, что чешуи защищают внутренние части почки и от холода, но это неверно: в почке нет того внутреннего запаса тепла, который могли бы сохранить многослойные покровы. Экспериментально было показано, что температура зимой внутри почки яблони отличается от наружной не более чем на ±0,5°С (ночью температура внутри почки ниже, чем днем). Однако чешуи смягчают вредное влияние резких перепадов температур, особенно рано весной. В таких случаях говорят о закрытых почках, в отличие от открытых, или голых, лишенных специально видоизмененных чешуи. Хорошими примерами закрытых почек могут служить зимующие почки деревьев и кустарников (дуб, береза, липа, лещина, ольха, бузина, ива и др.) и некоторых многолетних трав (грушанка, копытень и др., рис. 117, 4, 10). Число почечных чешуи у разных видов различно: иногда их свыше 20 (дуб), иногда всего 2 (ивы) или даже одна (почки некоторых однодольных).

Рис. 117. Строение закрытых почек:
/— дуб, общий вид и продольный разрез; 2— то же у бузины; 3— липа, общий вид и поперечный разрез; 4— копытень, общий вид растения (а), зимующая почка (б) и развернутые части почки (в); 5 — вишня, продольный разрез; 6— ясень, общий вид и продольный разрез; 7— черемуха; 8— тополь, общий вид и поперечный разрез; 9— бук; 10— грушанка; /, 3, 6— вегетативные почки; 2 и 4— вегетативно-генеративные почки; 5— генеративная почка; ПЧ — почечные чешуи; Л — зачатки листьев; Цв — зачаток цветка.
Открытую почку на верхушке имеют в течение весны и лета растущие побеги тех деревьев и кустарников, которые зимой имели закрытые почки, а также побеги многих однолетних и многолетних трав (рис. 118). Некоторые из травянистых многолетников зимуют с открытыми почками (живучка, зеленчук, кошачья лапка и др.).

Рис. 118. Строение открытых почек:
/— зимующие почки калины - гордовины; 2— береза, кончик растущего побега (а) и его верхушечная почка (б); 3—клевер, верхушка побега (а) и схема внутреннего строения почки (б); 4—побег злака; 5—схема продольного разреза его верхушечной почки, вегетативной (а) и вегетативно-генеративной (б).
Открытые почки (без специальных чешуи) свойственны также многим (но далеко не всем, как иногда думают) древесным породам влажных тропиков и субтропиков (в частности, цитрусовым).
Конус нарастания и самые молодые листовые зачатки в открытых почках, конечно, не «открыты» в буквальном смысле: они окружены, а часто и плотно закрыты частями более взрослых листьев. Так, у клевера каждый последующий зачаток листа полностью одет прилистниками предыдущего листа (рис. 118, 3). У растущего побега березы прилистники плотно окутывают не развернувшуюся еще пластинку «своего» листа (рис. 118, 2). У злаков верхушечная почка растущего побега заключена в трубку из влагалищ одного или нескольких более взрослых листьев (рис. 118, 4, 5).
Листорасположение. Листорасположение, или филлотаксис (греч. филлон — лист; таксис — расположение, порядок),— порядок размещения листьев на оси побега, отражающий его радиальную симметрию. Различают несколько основных вариантов листорасположения.
Спиральное, или рассеянное, листорасположение (иначе еще называемое очередным) наблюдается, когда на каждом узле расположен один лист и основания последовательных листьев можно соединить условной спиральной линией, растянутой вдоль удлиненного стебля или почти плоскостной на широком укороченном стебле (рис. 122, /, 2; 123, 5). Эту линию называют основной генетической спи ралью, так как она отражает последовательность заложения листьев, их генезис.
Двурядное листорасположение, которое можно рас сматривать как частный случай спирального, отражает так называемую маятниковую симметрию деятельности апекса. При этом на каждом узле находится один лист, охватывающий широким основанием всю или почти всю окружность оси. Средние линии (медианы) всех листьев лежат в одной вертикальной плоскости (рис. 122, 3; 123, 4).
Мутовчатое листорасположение возникает, если на одном уровне закладывается несколько листовых почек, образующих общий узел. Нередко при более детальном изучении выясняется, что каждый лист мутовки имеет собственный узел, но они очень сближены (рис. 122, 4; 123, /).
Супротивное листорасположение — частный случай мутовчатого, когда на одном узле образуются два листа, точно друг против друга, так что их медианы лежат в одной вертикальной плоскости; чаще всего (хотя и не всегда) такое листорасположение бывает накрест супротивным (рис. 122, 5; 123, 3), т.е. плоскости медиан соседних пар листьев взаимно перпендикулярны.
 Порядок
заложения листовых зачатков на апексе
побега — наследственный
признак
каждого
вида, иногда характерный для рода и
даже
целого семейства растений; он зависит
от генетических факторов. На
расположение листьев могут оказывать
влияние внешние факторы, прежде всего
условия освещения, а также сила тяжести,
направление действия которой
зависит от направления роста самого
побега. Таким образом, окончательная
картина листорасположения может сильно
отличаться от первоначальной
и обычно приобретает ярко выраженный
приспособительный характер.
Листья располагаются так, что их
пластинки оказываются в
наиболее благоприятных в каждом
конкретном случае условиях освещения.
Порядок
заложения листовых зачатков на апексе
побега — наследственный
признак
каждого
вида, иногда характерный для рода и
даже
целого семейства растений; он зависит
от генетических факторов. На
расположение листьев могут оказывать
влияние внешние факторы, прежде всего
условия освещения, а также сила тяжести,
направление действия которой
зависит от направления роста самого
побега. Таким образом, окончательная
картина листорасположения может сильно
отличаться от первоначальной
и обычно приобретает ярко выраженный
приспособительный характер.
Листья располагаются так, что их
пластинки оказываются в
наиболее благоприятных в каждом
конкретном случае условиях освещения.
Рис. 122. Основные типы листорасположения:
/ — спиральное у дуба (молодой растущий побег, видны прилистники);
2— схема основной генетической спирали; 3— двурядное у гастерии
(а—вид растения сбоку, б—вид сверху, схема); 4—мутовчатое у
олеандра; 5— супротивное у сирени.
Основные видоизменения побега. Видоизменения побега связаны с выполнением функций вместилища запасных веществ, перенесения неблагоприятного времени года, вегетативного размножения и др. Наиболее распространены следующие видоизменения (рис. 76, 77Корневище — многолетняя часть побега, расположенная под землей и несколько похожая на корень. Однако у корневища отсутствует корневой чехлик и оно несет редуцированные листья в виде чешуек. Обычно на корневище образуются придаточные корни. Корневища бывают длинные и тонкие (пырей, осока, см. рис. 76, Г) или короткие и толстые (щавель, цикута, касатик (см. рис. 76, В). Растут они верхушкой, где имеется почка. Ветвятся корневища так же, как стебель. Ежегодно корневища образуют из верхушечной и пазушных почек надземные однолетние побеги. Старые части корневищ постепенно отмирают. Корневища свойственны многолетним травянистым растениям. Растения с горизонтально ветвящимися корневищами, формирующими много надземных побегов, быстро занимают большую площадь. Если это сорняки (пырей), то борьба с ними довольно затруднительна. Растения с длинными корневищами используются для закрепления песков (колосняк). В луговодстве злаки с длинными горизонтальными корневищами называют корневищными (полевица белая, мятлик луговой и др.), а с короткими и малоразвитыми корневищами— кустовыми (ежа, тимофеевка и др.).
Стеблевые клубни — утолщенные части стебля, вместилища запасных веществ. Они бывают надземными и подземными. Надземные клубни представляют собой утолщение главного стебля (кольраби, см. рис. 76,Д) или боковых побегов (тропические орхидеи). Они несут нормальные листья. Подземные клубни являются или утолщениями подсемядольного колена (цикламен) или подземных побегов — столонов (картофель, см. рис. 76, £). Листья на подземных клубнях редуцируются. В пазухах их находятся почки, называемые глазками.
Луковица представляет собой укороченный стебель (донце), несущий многочисленные, тесно сближенные листья. На верхушке донца находится почка. У многих растений (лук, тюльпан, гиацинт и др.) из этой почки развивается надземный стебель, а из боковой пазушной почки формируется новая луковица (см. рис. 76,Л). Наружные чешуйки луковицы в большинстве случаев сухие, пленчатые и выполняют защитную функцию, внутренние — мясистые. По форме луковицы бывают шаровидные, яйцевидные, сплющенные и т. д. Луковицы бывают не только подземными, но и надземными, развивающимися в соцветиях (чеснок, дикий лук) или на стеблях в пазухах листьев (некоторые лилии). Они очень мелкие и представляют собой видоизмененные вегетативные или цветочные почки, служащие для размножения.
Клубнелуковица внешне похожа на луковицу, но морфологически ближе к клубню, все листовые чешуйки у нее сухие, а запасные питательные вещества откладываются в стеблевой части — донце (шафран, гладиолус, см. рис. 76,Б).
Колючки образуются из укороченных побегов (яблоня, груша, терн, боярышник и др.) и иногда ветвятся (гледичия, цитрусовые).
Колючки особенно характерны для растений жарких сухих местообитаний. Они служат для защиты от поедания животными (см. рис. 92, Б).
Усики образуются из пазушных почек у некоторых лиан (виноград, тыквенные), служат для прикрепления к опоре (см. рис. 93, Л, 270).

Кладодии представлены плоскими листовидными стеблями, расположенные в пазухах редуцированных листьев. На них образуются также редуцированные чешуйчатые листья, в пазухах которых развиваются цветки (рис. 77), Они встречаются у представителей многих семейств, живущих преимущественно в засушливых районах (иглица, филлантус и др.).
Рис. 77. Кладодии
иглицы:
/ — кладодий,
2 — цветок, 3 —лист

Рис. 76. Видоизменения побега. А — луковица лука; Б — клубнелуковицы шафрана;
В — корневище касатика; Г — корневище пырея; Д — надземный стеблевой клубень
кольраби; Е — подземные клубни картофеля:
1— сухие чешуйки, 2— сочные чешуйки, 3 — почка, 4 — донце, 5 — придаточные корни, 6 — материнский клубень, 7 - молодые клубни, 8 — столоны
Приспособительные изменения в листорасположении. Листовая мозаика.
Как уже говорилось, в процессе роста побега его листорасположение может меняться. Благодаря неравномерности роста стебель может скручиваться вокруг собственной оси, нарушая правильность листовых расположений. При сохранении симметрии размещения оснований листьев по стеблю черешки этих листьев могут изгибаться, поворачивая пластинки так, что по их расположению уже не удается определить исходную формулу и диаграмму. Особенно яркое выражение такие смещения получают в форме листовой мозаики, наблюдаемой чаще всего на побегах древесных и травянистых растений, а также на вертикально растущих розеточных побегах. При этом пластинки всех листьев располагаются
горизонтально, листья не затеняют друг друга, а образуют единую плоскость, где нет просветов; более мелкие по размерам листья заполняют просветы между крупными. Хорошие примеры листовой мозаики можно видеть на горизонтальных ветвях в кроне липы, на побегах плюща (рис. 127), герани табака, подорожника и др.

Рис. 123. Листовая мозаика у плюща (ползучие побеги).
ОПОРНЫЕ ТОЧКИ:
Побег - основной орган высшего растения, он выполняет функцию воздушного питания и способен к разнообразным метаморфозам.
Филлотаксис (греч. филлон — лист; таксис — расположение, порядок),— порядок размещения листьев на оси побега.
Междоузлия -участки стебля между соседними узлами.
Узел - участок стебля на уровне отхождения листа или мутовки листьев.
Листовая мозаика-это когда пластинки всех листьев располагаются горизонтально и не затеняют друг друга.
Лист – боковой орган побега
В отличие от корня и стебля лист является боковым органом ограниченного роста, нарастающим не верхушкой, а при основании - у однодольных, или же рост идет по всей поверхности с одинаковой скоростью - у двудольныех. У многолетних одревесневающих растений это временный орган, периодически возникающий заново. У одревесневающих растений продолжительность жизни листьев явно уступает продолжительности жизни стебля. У травянистых однолетников продолжительность жизни листа и стебля приблизительно одинакова и чаще всего исчисляется 45—120 днями. У вечнозеленых растений каждый лист функционирует от 1 до 5 лет, а у некоторых хвойных, например, у тисса, пихты - до10 лет.
Части листа. Лист обычно подразделяют на части (рис. 78): листовая пластинка, че решок, прилистники — выросты основания черешка, влагалище — расширенная нижняячасть листа, охватывающая стебель, язычок— пленочка или волоски на границе между пластинкой листа и влагалищем, ушки — боковые выросты пластинки листа на границе с влагалищем. Однако лист какого-либо конкретного растения имеет лишь две-три части, редко только одну — листовую пластинку (сидячие листья). Наиболее часто встречаются такие сочетания:
1) листовая пластинка и черешок, иногда еще и прилистники, как у многих бобовых, розанных и др.;
2) листовая пластинка, черешок и влагалище, как у некоторых Сельдерейных;
3) листовая пластинка, влагалище, язычок, а иногда и ушки — у злаков.
Очень редко бывают исключения: у папоротника лист растет, как и стебель, верхушкой, у южноафриканской вельвичии лист — постоянный орган и имеет неограниченный рост.
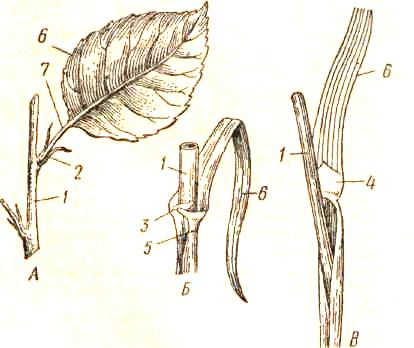
Рис. 78. Строение листа. А — яблони; Б — ячменя; В — злака.
1 — стебель, 2 — прилистники, 3 — ушки, 4 — язычок, 5 — влагалище, 6 — листовая пластинка, 7 — черешок.

Рис.
80. Переход от низовых листьев (защитных
чешуек) к срединным в почке красной
смородины;
чешуйки в данном случае представляют
листья с очень расширенным черешком
и
недоразвитой пластинкой, превращенной
в короткое
остроконечие
Функция черешков более определенна. Они ориентируют листовые пластинки по отношению к источнику света. Черешкам принадлежит важная роль в создании листовой мозаики, т. е. такого размещения листьев на побеге, чтобы они не затеняли друг друга (рис.123). Это достигается:
а) различной длиной и изогнутостью черешков;
б) различной величиной листовых пластинок;
в) разной светочувствительностью листьев.
Величина листьев сильно варьирует, нередко даже у одного индивидуума. В нашей флоре многие виды имеют очень мелкие листья величиной до 1 —1,5 мм. Особенно крупные листья у видов растений, преимущественно тропической и субтропической флор, например, у виктории (Южная Америка) плавающий лист имеет диаметр до 1,5 м. У некоторых пальм, например у африканской винной пальмы рафии, лист достигает 15 м в длину.
Формации листьев, гетерофиллия. Из трех основных органов растения лист наиболее изменчив по форме, величине, окраске, продолжительности жизни и др. Даже на одном растении в любой момент можно наблюдать неодинаковые листья.
Различают три формации листьев: низовую, срединную, верхушечную. Формацию низовых листьев составляют обычно недоразвитые листья или видоизмененные в связи с выполнением специализированной функции, например защитной или запасающей (рис. 80). Сюда относят: семядольные листья, кроющие чешуйки разнообразных почек, чешуйки (редуцированные листья) корневищ и надземных побегов. Срединные листья — это листья, типичные для данного вида. Они составляют основную массу, иногда довольно разнообразную по форме и величине. В отличие от предыдущих они всегда хлорофиллоносные. К формации верховых относят листья, расположенные в соцветиях, — прицветники, листовые обвертки. Это, как правило, недоразвитые листья, лишенные черешков; иногда они окрашены, иногда бесцветны. Они почти целиком утратили функцию фотосинтеза.
Когда говорят вообще о листьях конкретного растения, то имеют в виду именно срединную формацию листьев.


Рис. 82. Классификация простых листьев с цельной пластинкой (схема). А — по форме пластинки; Б — по форме верхушки; В — по форме основания; Г — по форме края:
1 — округлый, 2— овальный, 3 — продолговатый, 4 — линейный, 5 — яйцевидный, 6 — ланцетный, 7 — обратнояйцевидный, 8 — обратноланцетный, 9 — тупой, 10 — острый, 11 — заостренный, 12 —остроконечный, 13 — клиновидный, 14 — закругленный, 15 — сердцевидный, 16 — стреловидный, 17 — копьевидный, 18 — пильчатый, 19 — двоякопильчатый, 20 — зубчатый, 21 — колючезубчатый, или
шиповатый, 22 — городчатый, 23 — выемчатый
1
1
Иногда в структуре срединных листьев наблюдаются заметные различия, вызванные возрастными особенностями. Часто решающее влияние оказывает среда. Особенно контрастные различия наблюдаются у некоторых водных растений: лютика водяного, лимкофилы, стрелолиста, кабомбы (растение водоемов тропической Америки; у нас разводят в аквариумах) и др. Формация срединных листьев у них легко подразделяется на две-три зоны, резко обособленные морфологически: подводных, плавающих и надводных листьев. Это явление получило название гетерофиллии (рис. 81).
Простые и сложные листья. Простые листья составляют основное разнообразие и явно преобладают в мире растений. Они свойственны почти всем травянистым растениям и подавляющему большинству деревьев и кустарников.
Простые листья характеризуются:а) одной лишь листовой пластинкой, иногда рассеченной настолько глубоко, что создается впечатление большого числа пластинок (например, у полыни, петрушки и др.); б) простые листья или вовсе не опадают (большинство травянистых растений), или же имеют только одно сочленение между черешком и стеблем и опадают целиком (деревья и кустарники). Простые листья классифицируют по целому ряду признаков. Приведем лишь некоторые из них.
Листья с цельной пластинкой:
а) по форме листовой пластинки — округлые, овальные, яйцевидные, продолговатые, ланцетные, линейные, мечевидные, игольчатые и др. (рис. 82);
б) по характеру основания листовой пластинки — сердцевидные, округлые, клиновидные, стреловидные, копьевидные, низбегающие;
в) по характеру верхушки — тупые, острые, заостренные, остроконечные, выемчатые;
г) по характеру края листа — цельнокрайние, зубчатые, пильчатые, городчатые, выемчатые, двоякопильчатые, извилистые.
Рис. 83. Классификация
простых листьев с расчлененной
пластинкой (схема). А—лопастные; Б
— раздельные; В — рассеченные: / —
тройчато-, 2 —пальчато-, 3 — перисто-
Листья с расчлененной пластинкой:
а) лопастные— выемки достигают не более четверти ширины листовой пластинки (дуб, хлопчатник);
б) раздельные — выемки составляют одну треть пластинки и более (мак);
в) рассеченные — выемки достигают главной жилки листа (картофель).
В зависимости от взаиморасположения выемок и глубины надреза различают пальчатолопастные, пальчатораздельные, пальчаторассеченные; перистолопастные, перистораздельные, перисторассеченные листья (рис. 83).
Сложные листья свойственны только некоторым семействам, в частности семейству Бобовые, причем в типичном виде они представлены у деревьев и кустарников. В отличие
от простых сложные листья характеризуются несколькими, четко обособленными листовыми пластинками (листочками), каждый из которых имеет сочленение черешка листочка с общим черешком (рахисом). Между рахисом и стеблем есть еще одно сочленение. Таким образом, при листопаде отдельно опадают части сложного листа —листочки, отдельно — рахис. Среди травянистых растений также известны отдельные представители со сложными листьями, например, донники, люцерны и др.
Группу сложных листьев в зависимости от расположения листочков на рахисе подразделяют на две подгруппы: перистосложные и пальчатосложные (рис. 84).
П еристосложные:
еристосложные:
а) непарноперистосложные, когда верхушка рахиса заканчивается одним (непарным) листочком,
б) парноперистосложные, когда на верхушке рахиса два листочка;
в) тройчатые, когда сложный лист имеет только три листочка.
Перистосложные листья иногда бывают более сложной конструкции — дваждыперистсложные, как у настоящих акаций и др. Встречаются и многократно перистосложные листья.
Пальчатосложные: листочки располагаются не по длине рахиса, а лишь на верхушке и расходятся более или менее радиально.
Формы жилкования. Различают следующие формы жилкования (рис. 85).
Простое.Листовую пластинку пронизывает от основания до верхушки только одна жилка (проводящий пучок).Простое жилкование встречается у высших споровых: мхов, плаунов, у многих голосеменных (хвойных), а также у некоторых покрытосеменных(элодея).
Дихотомическое. Листовую пластинку пронизывает несколько вильчато разветвленных жилок (гинкго).
Рис. 84. Сложные листья. А — пальчатосложные Б — перистосложные: / — пальчатосложный конского каштана, 2 — тройчатый лимона трехлисточкого, 3-парноперистый чилиги, 4— непарноперистый шиповника, 5-двоякоперистосложный гледичии
Жилкование листьев. Сетчатое. Это наиболее широко распространенная форма жилкования. Из черенка в листовую пластинку входит одна жилка (первого порядка), которая затем дает ответвления — боковые жилки, образующие в своей совокупности очень густую сеть — перистое жилкование. У листьев перисторазрезанных будет соответственно и перистое жилкование.
Другой вариант сетчатого жилкования — пальчатое. В этом случае из черешка в листовую пластинку входят несколько жилок первого порядка, дающих сеть мелких разветвлений
Дуговидное и параллельное. Листовую пластинку от основания до верхушки пронизывает несколько неветвящихся одинаковых жилок. В одних случаях они расположены строго параллельно (например, у злаков, осок), в других — дугообразно (например, у ландыша).
Наиболее примитивной формой листа у голосеменных и покрытосеменных является простой лист, цельный или лопастной. Сильно рассеченные и сложные листья возникли позднее. Прилистники также рассматриваются как признак низкой организации.
Наиболее примитивным типом жилкования следует считать простое и дихотомическое.
Приспособительные изменения в листорасположении. Листовая мозаика.
Как уже говорилось, в процессе роста побега его листорасположение может меняться. Благодаря неравномерности роста стебель может скручиваться вокруг собственной оси, нарушая правильность листовых расположений. При сохранении симметрии размещения оснований листьев по стеблю черешки этих листьев могут изгибаться, поворачивая пластинки так, что по их расположению уже не удается определить исходную формулу и диаграмму. Особенно яркое выражение такие смещения получают в форме листовой мозаики, наблюдаемой чаще всего на побегах древесных и травянистых растений, а также на вертикально растущих розеточных побегах. При этом пластинки всех листьев располагаются
горизонтально, листья не затеняют друг друга, а образуют единую плоскость, где нет просветов; более мелкие по размерам листья заполняют просветы между крупными. Хорошие примеры листовой мозаики можно видеть на горизонтальных ветвях в кроне липы, на побегах плюща (рис. 127), герани табака, подорожника и др.
Рис. 123. Листовая мозаика у плюща (ползучие побеги).
ЦВЕТОК
Цветок представляет собой видоизмененный укороченный побег, приспособленный для образования микро- и мегаспор, гамет и для перекрестного опыления. В результате опыления и последующего полового процесса (оплодотворения) развиваются семена и плоды. Цветок имеют только покрытосеменные растения.
Типичный цветок покрытосеменных заканчивает собой главный или боковой побеги. В морфологическом отношении цветок является укороченным неразветвленным побегом. Стеблевая часть цветка представлена цветоножкой и цветоложем (рис. 94). У разных родовых групп растений цветоложе может приобретать разнообразную форму (рис. 95): от конической до плоской и даже вогнутой в виде бокала. На цветоложе размещены, чаще всего кругами (мутовками), реже по спирали, видоизмененные листья — цветолистики: чашелистики, лепестки, тычинки, пестики. В большинстве случаев цветок имеет 5 (или 4) мутовок (циклический цветок):,чашелистики —1 круг (чашечка), лепестки — 1 круг (венчик), тычинки — 1 или 2 круга (андроцей), пестики — 1 круг (гинецей). При спиральном расположении цветолистиков (ациклический цветок) число каждой их категории обычно неопределенное. Промежуточное положение занимают цветки гемициклические, у которых мутовчатое расположение одних элементов цветка сочетается со спиральным других (два круга околоцветника, множество тычинок и пестиков—по спирали).
Формула цветка Характеристику цветка можно давать сокращенно, в виде формулы где учитывается симметрия цветка, число кругов в цветке, число членов в каждом круге, срастание частей цветка и положение пестиков (верхняя или нижняя завязь). Части цветка обозначаются так: Р (Perigonium) -перигон (греч. пери-кругом, гони- колено) - простой околоцветник, К (Kelch) - чашечка, С (corolla) - венчик, А ( Androecenium )-андроцей, тычинки, G (Gynoecenium)-пестик (гинецей). Типы цветков имеют условные обозначения: ♂ ♀-обоеполый, -♀
пестичный, ♂ - тычиночный, - актиноморфный, или- зигоморфный.
Число членов отдельных частей выражаются цифрами: 5- лепестный венчик - Со 5, 6 членный андроцей – А6.
В случае срастания между собой цветолистиков число, указывающее на их количество, заключается в скобки, например, сросшийся 5 лепестный венчик -Со(5) или двубратственный андроцей - А(9)+1. Если отдельные доли чашечки расположены несколькими мутовками, то цифры, указывающие на количество их в отдельных мутовках, соединяются знаком +.
 Диаграмма.
Еще более полное представление о
строении цветка дает диаграмма, которая
является проекцией цветка на плоскость,
перпендикулярную его оси. Диаграммы
цветка демонстрируют расположение
частей цветка по отношению друг к другу
- ось соцветия вверху, а кроющий лист
внизу. Все члены цветка обозначают
всегда строго определенными значками.
Чашелистики обозначаются скобкой с
килем на спинке, лепестки - круглой
скобкой, тычинки - поперечным разрезом
через пыльник, для гинецея - поперечный
разрез через завязь. Диаграмма также
отображает срастание органов и
дополнительные детали цветка: нектарники
и др. . В случае срастания между собой
отдельных частей цветка знаки,
обозначающие их на диаграмме, соединяются
дугами или прямыми линиями.
Диаграмма.
Еще более полное представление о
строении цветка дает диаграмма, которая
является проекцией цветка на плоскость,
перпендикулярную его оси. Диаграммы
цветка демонстрируют расположение
частей цветка по отношению друг к другу
- ось соцветия вверху, а кроющий лист
внизу. Все члены цветка обозначают
всегда строго определенными значками.
Чашелистики обозначаются скобкой с
килем на спинке, лепестки - круглой
скобкой, тычинки - поперечным разрезом
через пыльник, для гинецея - поперечный
разрез через завязь. Диаграмма также
отображает срастание органов и
дополнительные детали цветка: нектарники
и др. . В случае срастания между собой
отдельных частей цветка знаки,
обозначающие их на диаграмме, соединяются
дугами или прямыми линиями.
Рис 124.Диаграмма цветка крестоцветного
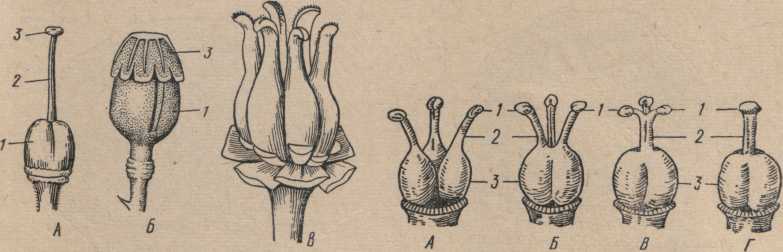
Околоцветник. Чашечка и венчик вместе составляют покров цветка, или околоцветник. Околоцветник, дифференцированный на различно окрашенную чашечку и венчик, называется двойным. Околоцветник, окрашенный в один цвет, называется простым (рис. 96). Простой чашечковидный околоцветник окрашен обычно в зеленый цвет (свекла, щавель и др.). Простой венчиковидный околоцветник окрашен ярко (тюльпан, гречиха и др.).
Цветки, совершенно не имеющие околоцветника, называются голыми (ясень, белокрыльник и др.)-
Чашечка состоит из более или менее плотных листочков, обычно зеленых, именуемых чашелистиками. Иногда чашечка ярко окрашена. В последнем случае она выполняет или усиливает роль венчика в привлечении насекомых-опылителей (фуксия, гранат и др.).

Рис. 94. Строение цветка Рис. 95. Цветоложе. А — вогнутое у шиповника; Б - (схема)): плоское у пиона; В — выпуклое у лютика.
/ — цветоложе, 2 — чашелистик,.
3 — лепесток, 4 — тычинка, 5 — пестик


Рис.97. Лепестки. А — сидячий цельный — лютика едкого; Б — ноготковый — гвоздики:
1 — ноготок, 2 — отгиб, 3 — чешуйка, прикрывающая нектарную ямку
Венчик состоит из более или менее окрашенных лепестков, которые образуют чаще всего второй (иногда и третий) круг цветка.
Разнообразие венчиков исключительно велико. Лепестки могут срастаться между собой, хотя бы частично, или оставаться свободными. В связи с этим различают два типа венчиков: раздельнолепестные (шиповник, вишня) и сростнолепестные, или спайнолепестные (вьюнок).
Если в раздельнолепестном венчике лепесток к основанию сужен, как лист в черешок, то это лепесток ноготковый (сем. Гвоздичные, Капустные и др.). Если основание широкое, округлое, лепесток называется сидячим (сем. Лютиковые, Розанные и др.рис. 97). Нередко встречаются и промежуточные формы лепестков,
В сростнолепестных венчиках различают: сросшуюся часть лепестков — трубочку, не сросшуюся — отгиб и место перехода трубочки в отгиб — зев (рис. 97).
Венчики, как и чашечки, могут быть: актиноморфные, или правильные, когда через венчик можно провести две или больше плоскостей симметрии (рис98), и зигоморфные, или неправильные, когда через венчик можно провести только одну плоскость симметрии.
Среди зигоморфных сростнолепестных наиболее часто встречаются следующие венчики (рис. 99):
Двугубый — отгиб состоит из двух неравноценных частей: верхней губы и нижней (сем. Яснотковые, Норичниковые).
Язычковый — от трубки отходят сросшиеся лепестки, имеющие вид язычка (сем. Астровые).
Шпористый — лепестки образуют полый вырост, именуемый шпорцем (сем. Норичниковые).
Зигоморфные раздельнолепестные венчики встречаются у бобовых, фиалковых.
Актиноморфные спайнолепестные венчики по форме бывают: воронковидными, колокольчатыми, трубковидными и др. (рис. 98). Своеобразную форму представляет так называемый колпачковый околоцветник, когда лепестки срастаются верхушками (виноград).

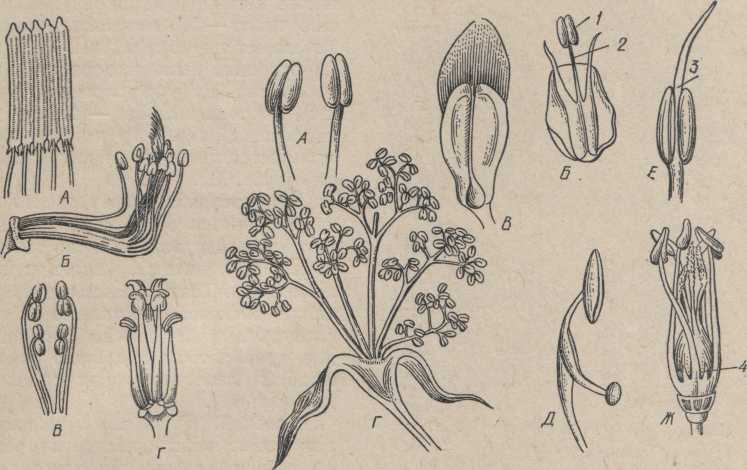
Рис. 102. Тычинки. А— розы морщинистой; Б — лука круглоголового; В — фиалки; Г — клещевины; Д — шалфея; Е — вороньего глаза; Ж — льна: 1— пыльник, 2 - тычиночная нить, 3 — связник, 4 — стаминодия
Связником, или спайником, называется обычно уплощенная, но нередко и утолщенная часть тычиночной нити, которая находится между обеими «половинками» пыльника. У пшеницы, ржи и др. он очень короткий, а у фиалки и у вороньего глаза, напротив, значительно удлинен и как бы пронизывает пыльник.
Пыльник обычно состоит из двух частей, или «половинок» — тек, расположенных по обе стороны связника. Каждая из тек в свою очередь состоит из двух гнезд, именуемых также пыльцевыми мешками, где развиваются микроспоры, а затем и пыльца (см. рис. 113). На разветвлениях сложных тычинок расположено только по одной теке (два пыльцевых мешка). Иногда и на простых тычинках развиваются только два пыль- цевых мешка (барбарис). Большое число — 50 пыльцевых мешков имеет тычинка омелы.
Строение пыльцы довольно однообразно, но морфология верхнего покрова пыльцы — экзины у различных семейств и порядков отличается широким разнообразием (рис. 103).
У некоторых видов растений часть тычинок остается недоразвитой и представлена лишь тычиночными нитями, лишенными пыльников. Такие бесплодные тычинки называются стаминодиями (аистник, лен). Иногда стаминодии приобретают вид ярко окрашенных лепестковидных пластинок (канна).
Рис.
103. Пыльцевые зерна. А — однобороздчатые
у магнолии крупноцветной (вверху) и
сусака зонтичного, (внизу); Б —
трехбороздчатые у джузгуна (вверху)
и пиона (внизу); В — многопоровое у
лютика азиатского; Г—многобороздчатое
у истода хохлатого
Гинецеем называют всю совокупность пестиков цветка, образованных плодолистиками (мегаспорофиллами).
Гинецей, который состоит лишь из одного пестика, называется простым, или одночленным. Гинецей, состоящий из двух или нескольких пестиков, называется сложным, или соответственно дву-, многочленным (см. рис. 95, 104, В). Пестик, состоящий из одного плодолистика, называется апокарпным. Сложный гинецей всегда апокарпный, ибо каждый пестик, его слагающий, состоит из одного плодолистика. Простой гинецей может быть образован одним плодолистиком - апокарпный, либо несколькими сросшимися плодолистиками — ценокарпный( рис 105).
Способы срастания плодолистиков ценокарпного гинецея могут быть различными (рис. 106).
Пестик — закрытое вместилище для семязачатков (семяпочек, или мегаспорангиев), образованное вследствие срастания одного или нескольких плодолистиков. Пестик состоит из завязи — вместилища семязачатков, столбика (одного или нескольких) и рыльца — приемника пыльцы (см. рис. 104). Иногда столбик отсутствует, тогда рыльце сидит как бы непосредственно на завязи — сидячее рыльце (мак). Различают завязь верхнюю и нижнюю в зависимости от положения ее по отношению к другим частям цветка. Верхняя завязь располагается
Б— простой мака; В — сложный сусака:
/ — завязь, 2 — столбик, 3 — рыльце
Рис. 105. Гинецей из трех плодолистиков (схема). А — сложный апокарпный, Б— Г — простой ценокарпный с разной степенью срастания плодолистиков:
/ — рыльце, 2 — столбик, 3 — завязь
свободно на плоском, выпуклом или вогнутом цветоложе и образуется только плодолистиками (см. рис. 95). В образовании нижней завязи, кроме плодолистиков, принимают участие и другие части цветка: цветоложе, основания чашелистиков, лепестков и тычинок, с которыми она срастается (рис. 107, В). Выделяют еще полунижнюю завязь, когда, по крайней мере, нижняя ее половина срастается с цветоложем или другими частями цветка.
В зависимости от числа не сообщающихся между собой гнезд завязи бывают одногнездными, двугнездными или многогнездными (см. рис. 106). Если гнёзда так или иначе сообщаются между собой, как у мака, завязь считают одногнездной.
Место прикрепления семязачатка к стенке завязи называется семяносцем, или плацентой. Семязачаток прикрепляется к плаценте семяножкой, или фуникулусом (см. рис. 106).
Около 75% видов покрытосеменных содержат в одном цветке и тычинки и пестики (обоеполые, или гермафродитные цветки). Около 25% имеют раздельнополые цветки — тычиночные и пестичные. Различают виды растений, у которых на одном экземпляре развиваются и тычиночные — «мужские» цветки, и пестичные — «женские» (кукуруза). Такие растения называют однодомными. Виды растений, у которых одни индивидуумы несут только тычиночные цветки, а другие — только пестичные, называют двудомными (конопля, тополь).
Соцветия. Цветки, как правило, собраны в соцветия, хотя у некоторых растений встречаются и одиночные цветки (роза, пион и т. д.). Под соцветием понимают побег, который несет цветки и видоизмененные вегетативные листья - прицветники (лист, из пазухи которого выходит цветоножка) и прицветнички ( листья , один или два, расположенные на цветоножке).
По способу ветвления различают два основных вида соцветий: моноподиальные и симподиальные.
В моноподиалъном соцветии четко выражена ось первого порядка. Развитие цветков осуществляется от основания к вершине. Различают простые и сложные моноподиальные соцветия. К простым моноподиальным соцветиям относятся:
колос - цветки лишены цветоножек и сидят на оси первого порядка,
початок - колос с сильно утолщенной осью. Початок окружен одним или несколькими листьями - чехол или крыло,
кисть - цветки имеют, как правило, цветоножки одинаковой длины,
щиток - кисть, у которой нижние цветоножки длиннее верхних и цветки располагаются в одной плоскости,
зонтик - главная ось укорочена, а цветоножки почти одинаковой длины и выходят как бы из одной плоскости,
головка - укороченная, булавовидная ось, цветоножек нет или они очень короткие,
корзинка - плотно сомкнутые цветки на сильно расширенном общем цветоложе верхушечные листья скручены и образуют обвертку.

Рис. 106. Простые моноподиальные соцветия; А- колос вербены, Б – початок белокрыльника, В – кисть черемухи, Г – щиток груши, Д – зонтик лука, Е – головка клевера, Ж – корзинка ноготка.
Моноподиальное соцветие называют сложным, когда ось первого порядка не несет отдельные цветки, а простые соцветия. К моноподиальным сложным соцветиям относятся:
сложный колос - на главной оси сидят простые колоски,
сложный зонтик - на оси первого порядка сидят простые зонтики.
метелка - соцветие, главная ось которого несет боковые ветвящиеся оси, заканчивающиеся цветками, имеет пирамидальное очертание и напоминает сложную кисть,
сложный щиток - соцветие, главная ось которого несет боковые ветвящиеся оси, заканчивающиеся простыми щитками.
Симподиальные соцветия не имеют выраженной главной оси цветения. Нередко развивается ложная ось, состоящая из осей различных порядков. Распускание происходит от верхушки к боковым ветвям. Когда цветки расположены в одной плоскости, зацветание идет от центра к переферии
Однолучевой верхоцветник ( монохазий)- в верхней части отходит только одна ось второго порядка, под цветком, расположенным на оси второго порядка, образуется ось третьего порядка .В зависимости от того како отходят подцветочные ветви в одну или разные стороны различают улитку, или завиток ( подцветочные ветви развиты только в одну сторону) или извилину подцветочные ветви отходят то в одну, то в другую стороны.

Рис. 111. Симподиальные соцветия. А — монохазий (завиток) окопника; Б—дихазий ясколки; В — плейохазий молочая
Дихазий- двулучевой верхоцветник, ось каждого порядка дает по две подцветочные супротивные оси, порядок повторяется несколько раз.
Плейохазий - многолучевой верхоцветник, или ложный зонтик, где от главной оси отходит несколько подцветочных осей, которые образуют мутовку и заканчиваются цветками.
К укороченным многолучевмкам относят ложную мутовку и полумутовку и пучок. У ложной мутовки цветки расположены вокруг стебля в одной плоскости более или менее замкнутое кольцо, если оно разорвано, то это полумутовка. Пучок образуется, когда цветоносные ветви коротки, а соцветие более или менее сжато в одной плоскости.
ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ
Опыление. Суть опыления состоит в переносе пыльцы из пыльников, где она формируется, на рыльце пестика. Различают два способа опыления: самоопыление и перекрестное опыление
Самоопыление может осуществляться только в обоеполых цветках и наблюдается только у нераскрывающихся цветков (у фиалок, льнянок, овсяниц, арахиса, ячменя ).
В ходе эволюции перекрестное опыление оказалось более прогрессивным. При таком опылении потомство может совмещать наследственные свойства обоих родителей, а это широкие возможности в борьбе за существование. Способы переноса пыльцы весьма разнообразны: ветром, водой, насекомыми, птицами, млекопитающимися и т. д. Для этого у цветков имеются приспособления для своих опылителей.
Различают две формы перекрестного опыления: гейтоногамия ( греч « гейтон»-сосед, « гамео» - вступить в брак) соседнее опыление, когда оно осуществляется в пределах одного растения, ксеногамия ( греч. « ксенос» - чужой, « гамео» - вступить в брак) - собственно перекрестное, когда пыльца с цветка одной особи переносится на рыльце цветка другой особи.
Для осуществления перекрестного опыления у цветов появились сложные и оригинальные приспособления дихогамия и гетеростилия.
Дихогамия - ( греч. «дихе» - на две части, « гамео» - вступать в брак) разновременное созревание тычинок и пестиков. Она выражена в двух формах: протандрия (греч. « протерос» - ранний, « андрос» - мужчина), когда тычинки созревают раньше пестиков ( бобовые, мальвовые, сельдерейные ); протерогиния ( греч « протерос» - ранний, « гине» - женщина), когда в обоеполом цветке пестики созревают раньше тычинок (злаковые, ситниковые, подорожниковые и др.).
Гетеростилия - рыльце пестика и пыльники тычинок расположены на разных уровнях. Это называется разностолбчатостью и , возможно, служит защитой от гейтоногамии, чем вернее достигается перекрестное опыление.
Энтомофилия. Переносчиками пыльцы чаще всего являются насекомые. Этот способ опыления называется энтомофилия ( греч. « энтомос» - насекомое ). Эволюция многих семейств цветковых и насекомых шла совместно и по пути узкой специализации. Если бы не было насекомых, то в природе не существовало бы такого разнообразия цветков. Иногда растение оказывается в полной зависимости от своего опылителя.
Энтомофильные растения обладают ярким венчиком или околоцветником и, часто, очень крупными цветками ( цветок раффлезии в диаметре 1 метр, кувшинки, тюльпаны, орхидные, пионы более 15 сантиметров в диаметре ). Многие энтомофильные растения имеют крупные соцветия (зонтичные, сложноцветные, мареновые и т. д.).
Еще большее значение имеют запахи цветков (запах сирени, фиалки, розы, ванили, имбиря, тухлой рыбы, навоза и пр.) многие насекомые путь к цветкам находят по запаху.
Некоторые насекомые поедают пыльцу. Пыльца очень богатый источник пищи, особенно белковой (15 -20 % белка). Некоторые насекомые поедают пыльцу, а другие кормят ей личинок.
Запах и окраска цветков является вторичными аттрактантами ( англ. Attract -привлекать ). Пыльца и нектар- первичными.
Зоофилия. В тропиках распространено опыление цветков различными позвоночными животными, особенно птицами и летучими мышами, посещающие цветы главным образом из- за нектара (колибри, цветочницы, нектарницы ). Для таких цветов характерны яркая окраска, обилие нектара, аромат отсутствует.
Для опыления цветков летучими мышами характерны ночное цветение, неприятный запах, крупные размеры, обильное выделение нектара, содержащего слизь, массовая продукция пыльцы и сильно выставленные из кроны соцветия.
Гидрофилия- вода играет роль опыляющего агента не только для растений, цветки которых погружены в воду, но и для таких, цветки которых расположены выше поверхности воды. У них пыльники часто нитевидной формы, тем самым повышается парение в воде. Пыльца тяжелее воды и не имеет экзины, предохраняющей содержимое от высыхания и женские цветки расположены ниже мужских, что позволяет пыльце случайно попасть на рыльце. У водяной лобелии цветки полностью перешли к самоопылению. Тычиночный аппарат изменен так, что попадание воды внутрь цветка исключено.
Анемофилия-( греч. анемос - ветер ). В полосе лесов умеренного климата приблизительно 20% всех видов анемофильны. Этот процент возрастает в полярных районах и снижается в тропиках. К ветроопыляемым относятся злаки, осоки, полыни, хмель, конопля, крапива, березы, осины, дубы, грецкий орех, платаны, вязы, орешник и др. Цветки их обычно мелкие и невзрачные, голые или с чашечковидным соцветием, лишенные запаха. Пыльца мелкая и ее много. У таких растений часто раздельнополые соцветия, пыльцевые нити длинные и волосистые перистые рыльца.
Автогамия- самоопыление. При этом осуществляется принцип « лучше самоопыление, чем никакого опыления». У многих растений вырабатываются специальные приспособления, резервирующие возможность автогамии. У некоторых растений цветки располагаются у поверхности почвы и приносят семена только благодаря самоопылению. Иногда самоопыление происходит случайно.
Оплодотворение.
Под оплодотворением понимают слияние двух половых клеток -мужской и женской гамет ( греч. гемете - супруга, геметес - супруг ). Мужская гамета у цветковых растений называется спермием, женская - яйцеклеткой. Процессу оплодотворения предшествует образование микро - и мега - спор, гамет и опыления.
Микроспорогенез. Этот процесс совершается в гнездах тычинок. Снаружи пыльник покрыт эпидермой. Под эпидермой находится фиброзный слой, представленный несколькими рядами крупных клеток без содержимого. Впоследствии этот слой способствует вскрытию пыльника. За фиброзным слоем идет дегенерирующий слой. Самый внутренний слой - выстилающий. Клетки его крупные, с густой цитоплазмой и несколькими ядрами.

Рис. 113. Пыльник (поперечный срез). А—одно пыльцевое
гнездо с археспорием; Б — вскрывшийся пыльник:
/ — эпидерма, 2 — фиброзный слой, 3 — дегенерирующий слой, 4 – клетки выстилающего слоя, 5 — археспорий (спорогенная ткань), 6 — пыльца
По мере созревания клетки этого слоя теряют свою форму и идут на питание развивающихся микроспор. Развиваются микроспоры из материнских клеток, которые называются архиспорием и находятся в пыльцевых гнездах. Диплоидные материнские клетки претерпевают редукционное деление. В результате получается четыре гаплоидных микроспоры. Каждая из микроспор содержит плотную цитоплазму и крупное ядро. Микроспоры обычно еще внутри пыльника прорастают и образуют пыльцу (пыльцевое зерно).

в
Рис. 114. Микроспора и ее прорастание. А — микроспора; Б — пыльцевое зерно; В, Г — формирование пыльцевой трубки:
1 — экзина, 2 — интина, 3 — вегетативная клетка, 4 — репродуктивная клетка,
5 — спермии.
Начало прорастания микроспоры связано с митотическим делением. В результате образуется маленькая репродуктивная клетка и большая вегетативная клетка. Снаружи пыльцевое зерно покрыто экзиной и внутренней оболочкой интиной. Экзина имеет несколько проростковых пор, прикрытых «пробочкой», которые могут растворяться в сахаристой жидкости рыльца. Интина эластична и лишена пор.
На рыльце пестика за счет вегетативной клетки развивается пыльцевая трубка. Она имеет форму тонкой нити иногда до 30 см в длину. В пыльцевой трубке за счет деления репродуктивного ядра образуются мужские гаметы - спермин. Пыльцевая трубка растет сквозь рыхлую ткань рыльца и столбика и проникает в семязачаток сквозь его покровы или через микропиле.
МЕГАСПОРОГЕНЕЗ И ЖЕНСКИЙ ГАМЕТОФИТ
Строение и типы семязачатков. Семязачаток состоит из нуцеллюса ( ядра), и ингументов (покровов) , края которых образуют узкий канал ( микропиле), через который пыльцевая трубка проникает к зародышевому мешку, а также фуникулюса (семяножки), посредством которого семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где нуцеллюс и интегументы сливаются, называют халазой.
Различают 5 основных типов семязачатков:
п р я м о й семязачаток. Микропиле находится на одной оси с рубчиком фуникулюсом. Семяножка очень короткая ( гречишные, ароидные).
о б р а щ е н н ы й семязачаток. Он повернут на 180 градусов, так что микропиле и рубчик рассоложены рядом, причем микропиле обращено к плаценте. Это самый распространенный тип семязачатков среди цветковых
п о л у п о в е р н у т ы й семязачаток. Нуцеллюс с интегументами повернуты на 90 градусов ( примуловые, норичниковые).
О д н о с т о р о н н е и з о г н у т ы й семязачаток. Характеризуется односторонним разрастанием нуцеллюса и интегументов. Встречается у бобовых и центросеменных.
Д в у с т о р о н н е и з о г н у т ы й семязачаток . Нуцеллюс имеет подковообразную форму. Встречается у отдельных представителей тех семейств, где обычно односторонне изогнутые семязачатки.
Количество семязачатков в завязи варьирует от одного до огромного количества.
В завязи Gymbidium из семейства орхидных обнаружено до 2 000 000 семязачатков.
Мегаспорогенез и мегагаметогенез осуществляется в семязачатках. Снаружи семязачаток покрыт ингументами, которые не смыкаются, оставляя отверстие - микропиле или пыльцевход. Конец семязачатка, противоположный микропиле, называется халазой. Под ингументами семязачатка расположен многоклеточный нуцеллюс, или ядро семязачатка. Вскоре после заложения семязачатка и развития нуцеллуса одна из его клеток начинает разрастаться - материнская клетка. Материнская диплоидная клетка приступает к мейозу. В результате мейоза образуется четыре гаплодных клетки. В дальнейшем три мегаспоры отмирают и только одна, обращенная к халазе, остается. Мегаспора всегда одноядерна и гаплоидна. Мегаспора сильно разрастается, ее ядро претерпевает три последовательных митотических деления. После первого деления образуются два ядра, которые перемещаются к двум противоположным полюсам прорастающей мегаспоры - зародышевого мешка, где осуществляется два последующих деления. В результате на каждом из полюсов образуется по четыре ядра. Все восемь ядер гаплоидны. Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится яйцеклеткой, две другие синергидами. В совокупности они формируют яйцевой аппарат. Три клетки на халазальном полюсе называются антиподами. Два оставшихся ядра перемещаются к центру и сливаются.

Рис. 115. Гинецей пролески. А — общий вид; Б — поперечный разрез завязи; В — продольный срез семязачатка:
1— завязь, 2 — столбик, 3 — рыльце, 4 — стенка завязи, 5 — гнезда завязи,
6 — семязачаток, 7 — семяножка, 8 — халаза, 9 — интегументы, 10 — микропиле,
Л — нуцеллус, 12 — зародышевый мешок, 13 — яйцеклетка, 14 — синергиды, 15 — антиподы, 16 — вторичное ядро
Так формируется вторичное ядро зародышевого мешка. После этого все компоненты зародышевого мешка готовы к оплодотворению.

Рис.116 Мегаспорогенез и метагаметогенез у покрытосеменных ( схема)
1-материнская клетка спор, 2-6 основные стадии развития зародышевого мешка, 7- зрелый зародышевый мешок.
ОПЛОДОТВОРЕНИЕ И РАЗВИТИЕ СЕМЕНИ
Развитие пыльцевой трубки. Пылинка, попав на рыльце пестика, прорастает. Содержимое пыльцы, одетой интиной, выпячивается через поры в экзине и образует пыльцевую трубку. Скорость роста пыльцевой трубки составляет 35 мм/час
Ядро клетки и генеративное ядро находятся на растущем конце пыльцевой трубки. Достигнув завязи пыльцевая трубка направляется к семязачатку и проникает в него через микропиле. Оболочка зародышевого мешка растворяется при соприкосновении с кончиком пыльцевой трубки. В зародышевом мешке пыльцевая трубка растет по направлению к яйцеклетке. Оболочка на кончике пыльцевой трубки разрывается и оттуда выходят два спермия. Один сливается с яйцеклеткой, а другой – с вторичным ядром зародышевого мешка или с одним из центральных ядер. Происходит двойное оплодотворение – характерная особенность покрытосеменных растений, не встречающаяся у голосеменных. Двойное оплодотворение открыл в 1898 г. Русский ботаник С. Г. Навашин.
Впоследствии из оплодотворенной яйцеклетки развивается зародыш, а из канальцевой клетки с оплодотворенным вторичным ядром – эндосперм. Эндосперм покрытосеменных триплоиден, что является принципиальным различием между покрытосеменными и голосеменными.
По форме спермии различны: палочковидные, червеобразные, способные к передвижению, несмотря на отсутствие жгутиков..
Формирование зародыша. Оплодотворенная яйцеклетка переходит в состояние покоя, зависящее от времени и внешних условий. Первое деление сопровождается заложением поперечной перегородки. Из терминальной клетки развивается зародыш, а из базальной клетки в дальнейшем образуется конус нарастания побега и семядоли. Первичный корень возникает из самой нижней клетки подвеска. У двудольных зародыш имеет две семядоли, подсемядольное колено, первичный корень, конус нарастания первичного побега ( иногда формируется зачаточные листочки или зачаточные почечки).
Формирование эндосперма. Эндосперм служит основным источником питательных веществ
У орхидных развитие эндосперма подавлено. Триплоидное ядро отмирает немедленно или через несколько делений.
Семязачаток превращается в семя. Кожура образуется из интегументов, отчасти из нуцеллюса, Стенка завязи образует околоплодник, окружающий семена, развивающиеся в завязи. Вся завязь превращается в плод.
СЕМЯ
В результате процесса двойного оплодотворения из семязачатка формируется семя. Семя состоит из зародыша и запасных питательных веществ, покрытых семенной кожурой. Зародыш семени развивается из зиготы, образованной в результате слияния спермия с яйцеклеткой. Клетка, возникшая в результате слияния другого спермия с вторичным ядром зародышевого мешка, развивается в питательную ткань семени — эндосперм. Синергиды и антиподы обычно дегенерируют и растворяются, интегументы превращаются в кожуру семени, а нуцеллус у большинства

Рис. 117. Семена покрытосеменных. А—с эндоспермом, который окружает зародыш (мак); Б — с эндоспермом, который лежит рядом с зародышем (пшеница мягкая); В — с запасными веществами, отложенными в семядолях зародыша (горох); Г — с эндоспермом, окружающим зародыш, и мощным периспермом (черный перец); Д — с периспермом, окруженным зародышем (куколь);
/ — семенная кожура, 2 — семенная кожура, сросшаяся с перикарпием, 3-перисперм, 4 — эндосперм, 5 - зародыш, 6 — почечка, 7 — корешок, 8 — семядоли
растений потребляется непосредственно в качестве питательного вещества при формировании зародыша семени, реже превращается в питательную ткань — перисперм.
Различают четыре типа семян в зависимости от того, где откладываются запасные вещества: в эндосперме, в перисперме, в зародыше, в эндосперме и перисперме (рис. 117).
Рассмотрим детальнее строение семени с эндоспермом на примере зерновки пшеницы, представляющей плод с одним семенем (рис. 118). Снаружи она покрыта довольно тонким пленчатым слоем, который трудно отделить от внутренней части зерновки. Этот слой является околоплодником, сросшимся с кожурой семени. Внутри расположены зародыш и эндосперм. Зародыш имеет сформированные вегетативные органы будущего растения: зародышевой корешок с корневым чехликом, корневое влагалище — колеоризу, зародышевый стебелек и почечку. В центре почечки хорошо заметен конус нарастания стебля, прикрытый зародышевыми листьями. Верхний зародышевый лист — колеоптиль — служит защитой для молодого проростка при прохождении через почву. Часть зародыша, примыкающая к эндосперму, называется щитком. Щиток — единственная развитая семядоля, выполняющая здесь функцию поглощения питательных веществ из эндосперма в период прорастания семени. На противоположной щитку стороне стебелька находится эпибласт — вероятно, остаток (рудимент) второй семядоли. Последний у многих злаков отсутствует. Эндосперм в периферической части (под семенной кожурой) имеет слой однородных клеток, содержащих алейроновые зерна,— алейроновый слой, в центральной части расположены крахмалоносные клетки.
Примером семени с запасными веществами в зародыше может служить семя фасоли (рис. 119). Снаружи оно покрыто довольно толстой семенной кожурой. На узкой вогнутой поверхности семени расположен рубчик — место прикрепления семени к семяножке. Здесь же находится микропиле (семявход). Роль его состоит в пропускании воды и газов внутрь семени. Непосредственно над микропиле помещается небольшой бугорок, образованный зародышевым корешком.


Рис 118. Зерновка пшеницы. А- схема продольно-поперечного разреза;
Б-зародыш на продольном разрезе: 1-волоски,2-околоплодник,3-кожура семени,4,5- эндосперм,6-щиток, 7-стебелек,8-почечка, 9листочки, 10-колеоптиль,11-эпибласт,12-корешок, 13-чехлик, 14-колеориза (корневое влагалище).
А

Рис. 119. Семя
фасоли. А—внешний вид; Б — зародыш:
1— зародышевый
корешок, 2 — микропиле, 3 — рубчик,
кожуры,4- семенной рубчик 5-остаток
кожуры,6-почечка, 7-семядоли
С наступлением благоприятных условий (температуры и влажности) семена всасывают воду и при достаточном доступе воздуха начинают прорастать, формируя проросток.
ПРОРАСТАНИЕ СЕМЯН. СТРОЕНИЕ ПРОРОСТКОВ
Хозяйственное значение семян. Значение семян для самих растений очевидно. Но семена также имеют жизненно важное значение в хозяйстве. Нельзя забывать, что именно из семян (точнее, плодов-зерновок) злаков (пшеницы, риса, кукурузы, различных просовых злаков) получается основной продукт питания человечества — хлеб и крупы. Почти такое же значение имеют семена различных бобовых (горох, соя и др.). Велика пищевая ценность семян различных «орехоплодных» (кокос, грецкий орех, фундук и др.). Из семян масличных растений (подсолнечник, лен, кунжут, некоторые пальмы и т.д.) добываются пищевые и технические растительные масла. Семена используются также как пряность (тмин, анис, черный перец и др.). Ясно поэтому, что урожай семян культурных растений должен быть во много раз больше, чем нужно для поддержания и возобновления численности самих растений.
Урожайность в значительной мере зависит от качества посевного материала. Для посева отбираются вполне зрелые, полноценные, самые лучшие семена. Один из важных показателей качества семян — их всхожесть.
Прорастание семян. Как известно, для прорастания семян необходимы определенные условия. Главные из них — наличие воды (ткани зрелых семян предельно обезвожены, содержат не более 10—15% влаги) и достаточный доступ воздуха, обеспечивающий процессы интенсивного дыхания прорастающих семян. Кроме того, для каждого вида растений существует определенная температура, ниже которой семена не могут начать прорастать. Оптимальные температуры, при которых прорастание идет наиболее активно, почти для всех растений лежат в пределах +25-35°С, тогда как минимальные сильно колеблются: у растений умеренных и холодных климатов они могут быть немногим выше нуля (красный клевер -0,5оС, рожь +1°С, лен +2ОС, пшеница +4°С), а у жителей субтропиков и влажных тропиков — от 10 до20°С и выше (рис -10°C, дыни, огурцы -15.-l8°C и т. д.). Для прорастания некоторых семян (сельдерей, барбарис) благоприятна переменная температура. Семена многих дикорастущих растений умеренных и холодных климатов не могут прорасти без промораживания. Свет влияет на прорастание семян неоднозначно. Семена многих растений безразличны к свету, семена других не могут прорасти без света. Легко прорастают на свету семена моркови, лугового мятлика. Только при воздействии света прорастают семена салата, табака; только в темноте — семена фацелии, некоторых вероник.
Покой семян. Однако не всегда семена прорастают даже при самых благоприятных внешних условиях. Часто наблюдается так называемый глубокий покой семян, причины которого могут быть различными. Одна из них — недоразвитость зародыша в зрелом семени, о чем уже говорилось выше. В таких семенах зародыш может довольно долго доразвиваться за счет эндосперма, прежде чем приобретет способность к выходу из семенной кожуры и дальнейшему росту.
Один из характерных примеров — женьшень, семена которого в природе прорастают только на 3-й год после опадения с материнского растения. Поэтому первые опыты посева женьшеня в культуре и были неудачными. Другие примеры семян такого типа приведены на рисунке. Но задержка в прорастании может быть вызвана не только незрелостью зародыша. Нередко покровы семени, только что отделившегося от материнского растения, бывают водонепроницаемыми (например, у многих бобовых), слишком твердыми и механически препятствующими прорастанию (например, у косточковых плодовых). Иногда в тканях околоплодника и семенной кожуры вырабатываются вещества-ингибиторы (лат. inhibitio — задержка, удерживание), химически тормозящие прорастание (у некоторых ясеней, ряда тропических деревьев). У многих злаков, табака, бальзаминов внутренние слои семенной кожуры обладают пониженной газопроницаемостью. У растений умеренного климата эндогенный покой зародыша часто сочетается с непроницаемостью и твердостью кожуры. Покой семян, таким образом,— широко распространенное явление, которое можно рассматривать как важное приспособительное свойство, выработавшееся у цветковых растений в процессе эволюции. Благодаря покою семена предохраняются от преждевременного прорастания, что было бы особенно опасно в климатах с суровой зимой. В течение определенного времени покровы семян естественным образом разрушаются, перегнивают, ингибиторы прорастания из них вымываются, зародыши доразвиваются и т. д., и тогда семена получают возможность прорасти.
Типы семян по скорости прорастания и сохранению всхожести. Далеко не у всех растений семена впадают в длительный покой. Семена некоторых растений прорастают сразу и очень скоро теряют всхожесть; если они не смогли прорасти в течение немногих дней или недель, они погибают (например, у ив и тополей).
Можновыделить несколько групп растений по сочетанию признаков скорости прорастания и длительности сохранения всхожести.
Семена обладают длительным глубоким покоем и очень долго сохра няют всхожесть. Прорастают нередко через год, два и более после опадения. К этой группе относятся многие древесные растения и лесные травы. Опыты с монолитами луговых и полевых почв показали, что в почве сохраняются огромные запасы семян дикорастущих растений, в том числе полевых сорня ков; они могут прорастать порциями в течение десятков лет (в опытах — до 40—50 и даже 90 лет). Неудивительно поэтому, что борьба с многими сорняками так трудна. До 200—250 лет сохраняют всхожесть семена лотоса.
Семена прорастают сразу или вскоре после опадения (нередко после перезимовки) и сохраняют всхожесть в течение нескольких лет (7—12— 18). К этой группе принадлежит большинство культурных растений — зла ков, овощных — и многие дикорастущие луговые и степные травы.
Семена прорастают сразу и очень быстро теряют всхожесть. Сюда от носятся, кроме уже упомянутых ив и тополей, мать-и-мачеха клевер пол зучий, куколь, многие растения влажных тропиков.
Семена прорастают прямо на материнском растении, до опадения. Это так называемые «живородящие» растения, примеры которых очень немногочисленны. Гораздо чаще встречаются растения, которые совсем не дают плодов и семян, но их цветки метаморфизируются в маленькие по беги, опадающие и прорастающие в новые особи. Такие растения тоже называют живородящими, но это не настоящее живорождение. Данное явление характерно для крайних условий жизни: тундры, высокогорья, пус тыни (см. гл. VI).
Один из наиболее известных случаев — виды Rhizophora и Avicennia — растений мангровых зарослей, образующихся в экваториальной зоне по берегам океанов в полосе прилива и отлива (рис. 83). У них из завязавшихся плодов высовывается и начинает быстро расти мощный гипокотиль проростка с зачатком корня на конце. Достигая иногда 50—70 см и имея значительную массу, такой проросток отрывается от материнского растения и падает корнем вниз в илистую зыбкую почву, где сразу можно «заякориться» и не быть смытым очередным приливом. Естественно, что такое приспособление могло развиться только в особых почвенных условиях и во влажном тропическом климате.
Рис.
83. Прорастание семян ризофоры Rhizophora
mangle
на материнском растении:
1— ветка с плодами;
2—
отделившийся проросток; 3—5—
последовательные
стадии прорастания; 6—
разрез завязавшегося
плода; 3 — зародыш;ГП
— гипокотиль;ГК
— главный
корень
(зачаток);СМ
— семядоли;ПЧ
— почечка
проростка. дновременно
с поглощением воды начинается
активная ферментативная деятельность,
приводящая к мобилизации
запасных веществ т. е.
переходу их в растворимое
состояние, доступное для
поглощения клетками меристемы
зародыша. В частности, крахмал
превращается в растворимые сахара.
Эти процессы обеспечиваются
энергией за счет очень интенсивного
дыхания семян.
дновременно
с поглощением воды начинается
активная ферментативная деятельность,
приводящая к мобилизации
запасных веществ т. е.
переходу их в растворимое
состояние, доступное для
поглощения клетками меристемы
зародыша. В частности, крахмал
превращается в растворимые сахара.
Эти процессы обеспечиваются
энергией за счет очень интенсивного
дыхания семян.
Обычно семядоли зародыша выделяют ферменты, которые способствуют преобразованиям веществ эндосперма или перисперма. Таким образом, первая функция семядолей в семенах с внезародышевыми запасающими тканями - выделительная,так сказать, пищеварительная. В дальнейшем функция всасывания питательных веществ зародышем осуществляется также преимущественно семядолями, имеющими с самого начала большую всасывающую или сильно увеличивающими эту поверхность перед прорастанием. Один из наиболее эффектных примеров огромного разрастания семядоли в качестве сосущего органа (гаустории) — зародыш и проросток кокосовой пальмы. На рисунке 84 показано, как по мере прорастания кокосового ореха недоразвитый вначале зародыш дает надземный побег и придаточные корни, а семядоля тем временем врастает все глубже в жидкую часть эндосперма (кокосовое молоко) и всасывает его, в конце концов заполняя всю полость ореха сплошной губчатой массой.
Надземное и подземное прорастание. В результате усиленного питания прежде всего начинают разрастаться все органы зародыша. Из разрыва кожуры или из микропилярного отверстия первым обычно выходит зародышевый корешок, укрепляющий молодое растение в почве и начинающий самостоятельно всасывать извне воду и минеральные вещества. Одновременно растет и гипокотиль, проталкивающий кончик корешка в почву.
Семядоли ведут себя по-разному в зависимости от своих первоначальных функции, от дальнейшего поведения гипокотиля и от распределения зон роста в самих семядолях. Если запасы питательных веществ находились вне зародыша, то, как уже говорилось, семядоли прежде всего всасывают
Пч
МЭ
1— кокосовый орех (односемянный плод) до прорастания; недоразвитый зародыш (3) окружен твердой частью эндосперма (ТЭ), внутри — жидкая часть эндосперма — «кокосовое молоко» (МЭ); Пл — околоплодник; 2—прорастающий орех; видно разрастание семядоли (См), выполняющей функции гаустории; ПК — придаточные корни; Пч — развертывающаяся почка.
эти вещества. Затем благодаря росту гипокотиля или своего собственного основания (у однодольных) они могут быть вынесены на поверхность почвы, позеленеть и стать первыми ассимилирующими органами проростка, переходящего, таким образом, на автотрофное питание. В этом случае говорят о надземном прорастании (рис. 85, /—3). В других случаях семядоли так и остаются под землей, ограничиваясь гаусториальной функцией, а первыми ассимилирующими органами становятся следующие за семядолями листья (рис. 85, /, 4, 5). Это так называемое подземное прорастание. Если запасы были сосредоточены в тканях самих семядолей (в безэндоспермных семенах), то они прежде всего отдают эти запасы меристематическим тканям зародыша, а сами либо сразу же после этого сморщиваются и отмирают, не выходя из семени, как у дуба, гороха (рис. 86, 12, 14), т. е. прорастание бывает подземным, либо, реже, все-таки выходят после этого на поверхность, зеленеют и функционируют некоторое время как ассимилирующие органы.
Рис.
85. Схема надземного и подземного
прорастания двудольных:
/—
начало прорастания семени; 2,3-этапы
надземного
прорастания; 4,
5— этапы
подземного прорастания;
См
—
семядоли; Эпк
—
эпикотиль; ГК
—
главный корень; БК
— боковые
корни; ПК
— придаточные
корни; Чш
—
чешуевидные листья.
Гипокотиль черного цвета.
Дальнейшее развитие проростка. Типы проростков. Как мы видели, в процессе прорастания, кроме увеличения размеров и смены функций зародышевых органов, происходит и новообразование органов. В почечке на конусе нарастания продолжают закладываться листовые зачатки, в то время как ранее заложенные развертываются, а участки стебля между ними разрастаются, образуя междоузлия. Зародышевый корешок, превратившийся теперь в главный корень проростка, по мере роста ветвится; боковые корни вместе с главным образуют первичную систему главного корня. В области корневой шейки, на гипокотиле и в нижних узлах стебля могут рано образовываться придаточные стеблеродные корни.
Разнообразие облика проростков зависит не только от надземного или подземного положения семядолей, но и от особенностей роста главного побега и от соотношения между развитием системы главного корня и стеблеродных придаточных корней. Если все междоузлия заметно разрастаются, главный побег
становится удлиненным (рис. 86, /, 3, 5, 12, 13, 14). Иногда же рост междоузлий главного побега заторможен и они остаются укороченными. При этом узлы и листья сближены и побег выглядит как розеточный (рис. 86, 6—//) или укороченный побег с ложным стеблем, образованным лишь влагалищами листьев (что обычно для однодольных, в том числе для злаков. Система главного корня иногда развивается слабее, чем система придаточных корней, которые закладываются у некоторых злаков уже в зародыше. Получается так называемая мочковатая корневая система, характерная для проростков большинства однодольных.
Семядоли, как правило, имеют очень простую форму (круглые, овальные, продолговатые, без зубцов по краям; рис. 86, 1—3, 5—8, 14), хотя есть и исключения (рис. 86, 4). Первые листья могут быть, как уже упоминалось, недоразвитыми чешуевидными (при подземном прорастании) или зелеными ассимилирующими, но по форме и размерам они часто сильно отличаются от листьев взрослого растения данного вида: они мельче и проще по очертаниям. От таких юношеских листьев до типичных листьев взрослого побега можно проследить ряд постепенных изменений — листовую серию (рис. 86, 5, 8).
Таким образом, у проростков уже имеются все основные вегетативные органы — корневая и побеговая системы, которые продолжают в дальнейшем нарастать за счет верхушечных меристем и ветвления.

Рис. 86. Проростки двудольных:
1 — 11-е надземным прорастанием; 12—14—с подземным прорастанием: /—5 и 12—14— с удлиненным главным побегом; 6—11— с розеточным главным побегом; /(аи б) — фасоль обыкновенная; 2— тыква; 3 (а и б) — клещевина; 4— липа; 5— ясень обыкновенный; 6— копытень; 7— лютик ползучий; 8— костяника; 9— тмин; 10— редис; //— подорожник большой; 12— дуб черешчатый; 13— настурция; 14— горох.
Опорные точки:
Зародыш семени развивается из зиготы, образованной в результате слияния спермия с яйцеклеткой.
Клетка, возникшая в результате слияния другого спермия с вторичным ядром зародышевого мешка, развивается в питательную ткань семени — эндосперм.
Зародыш имеет сформированные вегетативные органы: зародышевый корешок с корневым чехликом, корневое влагалище — колеоризу, зародышевый стебелек и почечку.
Верхний зародышевый лист у однодольных— колеоптиль — служит защитой для молодого проростка.
Щиток у однодольных — единственная развитая семядоля.
Эпибласт —остаток (рудимент) второй семядоли у однодольных.
Вопросы для повторения
Чем отличаются семена однодольных и двудольных растений?
Что общего можно найти в строении семени однодольного и двудольного растения?
Что такое эндосперм и перисперм из чего они образуются?
Имеется ли колеоптиль у однодольных растений?
Из чего развивается эндосперм?
ПЛОД
Плод представляет собой орган размножения (а нередко и распространения), возникающий из цветка.
Плоды образуются в результате изменения гинецея, происходящего после двойного оплодотворения. У некоторых растений (виноград, цитрусовые и др.) видоизменению пестика не предшествует оплодотворение яйцеклетки. Такие плоды называют партенокарпическими, они обычно не содержат семян. В образовании плода, кроме гинецея, могут принимать участие прирастающие к нему части цветка (цветоложе, основания тычинок, лепестков, чашелистиков), а иногда и части соцветия. Наиболее существенная часть плода — заключенные в нем семена. Стенка плода называется околоплодником, или перикарпием.
Околоплодник формируется из стенок завязи и состоит из трех слоев: а) экзокарпия (наружный слой); б) мезокарпия (средний слой); в) эндокарпия (внутренний слой) (см. рис. 124).
Плод называют простым, если в образовании его принимает участие только один пестик. Иногда простые плоды могут распадаться по гнездам или разламываться на односеменные членики. Такие плоды называют дробными (см. рис. 121, Е, 122, Е). Плод называют сложным, или сборным, если он образован несколькими пестиками одного цветка рис. 121,5, 122,3, Я, 124,5).


Плоды, развивающиеся не из одного, а из нескольких цветков, так или иначе сросшихся между собой (свекла), или же из целого соцветия (шелковица, инжир, ананас и др.), называются соплодиями (рис. 120)-В образовании соплодий, кроме цветков, могут принимать участие и оси соцветия.
Разнообразие плодов исключительно велико. Оно вызвано главным образом приспособлением плодов к распространению. Поскольку еще нет естественной (филогенетической) системы плодов, которая могла бы широко применяться в практике определения растений, приходится придерживаться чисто морфологической классификации, в значительной мере искусственной. В основу этой классификации положены следующие признаки: 1) консистенция околоплодника (сухой или сочный); 2) число семян (много или одно); 3) вскрывание околоплодника (нераскрывающийся или раскрывающийся и способ раскрывания); 4) число плодолистиков, образующих плод.
ПЛОДЫ С СУХИМ ОКОЛОПЛОДНИКОМ
I. Коробочковидные (плоды многосеменные, обычно растрескивающиеся, рис. 121). 1.Листовка—одногнездный плод, образованный одним плодолистиком. Вскрывается по брюшному шву — линии срастания краев плодолистика. Из сложного гинецея развивается плод сложная листовка.
2.Боб — одногнездный плод, образованный одним плодолистиком. Вскрывается по брюшному и по спинному швам — двумя створками. Характерен для бобовых. Бобы могут иногда разламываться на отдель ные членики (копеечник, сераделла). Кроме типичных бобов, спирально закрученные (люцерна посевная), односеменные и тогда не вскрывающиеся (эспарцет).
3.Стручок, стручочек — двугнездный плод, образованный двумя плодолистиками. Семена прикрепляются к продольной перегородке. Вскрывается двумя швами. Стручочек отличается от стручка соотношением длины и ширины: если у стручка длина превышает ширину в 4 и более раза, то у стручочка длина превышает ширину не более, чем в 2—3 раза. Характерны для капустных. Стручки могут быть дробными.
4.Коробочка — плод, образованный двумя или несколькими плодолистиками. У разных растений различны способы вскрывания коробочки— дырочками, крышечкой, зубчиками, створками и т. д.
II ОРЕХОВИДНЫЕ ПЛОДЫ (плоды односемянные, при созревании не растрескиваются рис.123).
Орех, орешек — околоплодник жесткий, деревянистый.
Желудь — орех, у основания окруженный чашевидной плюской, развивающейся из расширения оси, несущей цветок.
Семянка — околоплодник кожистый, семя не срастается с около плодником. Из сложного гинецея образуется плод сложная семянка.
Крылатка — семянка, околоплодник которой имеет кожистый или перепончатый крыловидный вырост.
9.Зерновка - околоплодник кожистый, сросшийся с семенной кожурой. Характерна для злаковых
ПЛОДЫ С СОЧНЫМ ОКОЛОПЛОДНИКОМ
III. Ягодовидные (плоды с сочным эндокарпием, большей частью многосеменные, рис. 122):
10Ягода — весь околоплодник за исключением тонкой кожицы (экзокарпия) сочный, мясистый.
11. Яблоко — плод, в образовании которого, кроме завязи, принимает участие и цветоложе. Характерно для розанных (подсемейство Яблоневые).
12. Тыквина - плод, который развивается из нижней завязи, образованный тремя плодолистиками; экзокарпий жесткий, деревянистый. Характерна для тыквенных.
13. Померанец - экзокарпий окрашенный, с вместилищами, заполненными эфирным маслом; мезокарпий сухой, губчатый, белый; эндокарпий сочный, мясистый. Характерен для цитрусовых.
14.Костянка-околоплодник резко дифференцирован на экзо, мезо и эндокарпий .Косточка это деревянистый эндокарпий. Изредка встречается плод сухая костянка, например у миндаля, кокосовой пальмы.
Рис. 121. Коробочковидные
плоды. А — листовка морозника; Б —
сложная листовка водосбора; В — боб
гороха; Г — стручок капусты; Д —
стручочек ярутки; Е — дробный стручок
редьки; Ж — коробочка мака; 3 — коробочка
белены; И — коробочка дурмана

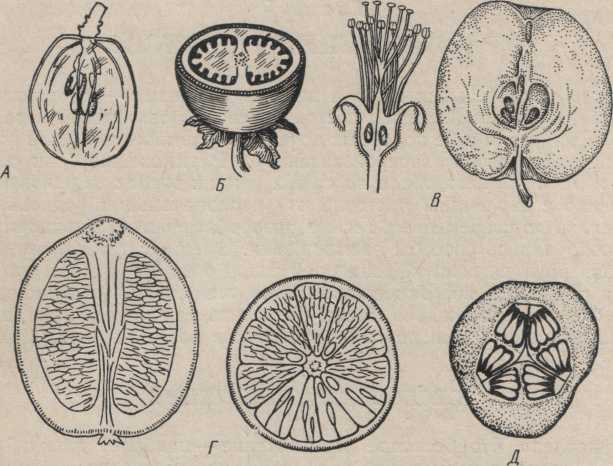

Рис. 123. Ореховидные плоды. А—орех лещины; Б — орешек гречихи; В — зерновка пшеницы; Г — желудь дуба; Д — крылатка вяза; Е — дробная крылатка клена; Ж — семянка подсолнечника (внешний вид и продольный разрез); 3 — сложная семянка лютика; И — сложная семянка земляники