

Аналитическая систематика — идиография
В область идиографии23 входят три задачи: 1) описание видов и других таксономических единиц; 2) разграничение видов и других таксономических единиц; 3) вопрос о таксономических единицах разного порядка.
1. Описание. Хотя в настоящем разделе мы касаемся практической систематики и задача описания как будто сужена практическими потребностями, не следует понимать практику в смысле голого определения. Слово «практический» имеет общеметодический смысл, и потому вопрос об описании надо поставить во всей широте, и прежде всего поставить вопрос: возможно ли полное описание, т. е. возможно ли охватить конечным числом слов все богатство признаков морфологических, эмбриологических, физиологических, химических и т. д., так как все такие признаки могут быть использованы как для самой систематики, так и для других целей. Обычный взгляд на организм как на агрегат независимо изменяющихся частей и признаков, конечно, не допускает возможности действительно полного описания. С этим взглядом стоит в связи и чисто генетическая трактовка системы, и такое частное мнение, как допущение существования
{53}
биологических видов и рас, т. е. таких, которые различаются исключительно физиологическими, экологическими и другими признаками при полном сходстве морфологических признаков. Напротив, взгляд Скиапарелли на организм как на чистую форму (т. е. такую, по любому, сколь угодно малому отрезку которой можно восстановить всю форму) даже в модифицированном отражении (принимая организм как комплекс чистых форм) допускает возможность полного описания. Поэтому сейчас ряд биологов выдвигают положение, что, может быть, число истинных признаков организма не бесконечно, а конечно. Этот взгляд получил свое выражение, например, в одной из работ П. В. Те-рентьева, который разделяет все возможные плеяды, связанные коррелятивной связью, и тогда достаточно будет от каждой плеяды знать один или несколько ведущих признаков, чтобы на основании их сконструировать весь организм. Не пытаясь решить этот вопрос (для этого необходимо проделать еще очень много конкретных исследований), можно сказать, что, во всяком случае, надо стремиться к тому, чтобы связывать ряды признаков в системы коррелятивных связей, и тогда описание приобретет гораздо большую полноту при гораздо большей краткости. Под ведущими признаками следует понимать такие, которые всего удобнее принимать как независимые при определении по ним иных зависимых признаков. При точном описании в отношении каждого признака надо определить не только его среднюю величину (арифметическую, геометрическую, медиану, моду), но и дисперсию, в общем случае построить кривую распределения. Вероятно, можно выставить такой биологический постулат, что, как бы ни были близки между собой две таксономические группы, нельзя найти ни одного признака, по которому они не отличались бы. Например, все млекопитающие имеют четыре конечности, лишь китообразные имеют редуцированные задние конечности. Не говоря уже о том, что в последнем случае, введя балл для характеристики развития задних конечностей, можно характеризовать животное дробным числом конечностей; даже в том случае, когда конечности имеют полное развитие, нельзя говорить о полном тождестве их числа для любых двух групп организмов; бывают млекопитающие уроды с редуцированным числом конечностей (описана даже родившаяся совершенно безногая овца). Склонность к таким уродствам различна у разных видов, и, беря генеральную совокупность особей двух соседних видов и изучая кривые распределения, мы, весьма возможно, найдем различие и по этому, казалось бы, совершенно стабильному признаку.
Очень важное понятие — кондициональные признаки (о них говорится и у Дарвина), на что обращал внимание В. Н. Беклемишев в лекциях по теории системы в Пермском университете. Группа может характеризоваться не тем или иным признаком, выраженным у всех своих индивидов, а определенной способностью проявлять тот или иной признак при каких-то, нам очень
{54}
часто вовсе неизвестных, условиях. Хороший пример таких кондициональных признаков — образование вдавления на брюшке у самцов ряда родов Gallerucinae (Phyllobrotica, Euluperus, Luperus; см. монографию Оглоблина в «Фауне СССР», с. 189, 191, 195, 201, 205, 279, 281). В разных родах это вдавление, несомненно, развивалось независимо, что можно заключить как из серии постепенного развития таких вдавлений в каждом роде, так и из отличительных родовых черт таких вдавлений. Кондициональным признаком для таких групп, как Orthoptera (Saltatoria), является способность образовывать совершенно между собой несходные стридуляционные органы; то же — стридуляционные органы совершенно различного характера— наблюдается у личинок в ряде родов пластинчатоусых жуков. Упомяну еще любопытный факт (видел целый атлас южноамериканских гусениц сфингид в Nature novitates, издание музея Ротшильда) — образование рисунка, напоминающего голову змеи на самых разнообразных местах тела (передний, задний конец и т. д.) гусениц сфингид. Можно указать и сравнительно редкий случай, где мы можем выставить и причину реализации кондиционального признака: при переходе членистоногих от водного образа жизни к наземному могут образоваться очень сходные органы: трахеи насекомых, сольпуг и мокриц.
Весьма возможно, что при дальнейшем углубленном изучении в связи с торжеством идеи о полифилетическом происхождении разных групп большинство признаков окажутся кондициональными. В пользу этого говорит уже одно то, что в самых естественных группах (что известно было и Дарвину) очень трудно найти признаки, характеризующие данную группу в целом, например ракообразные. Много примеров в отношении систематики блошек у Гейкертингера; из чтения книги С. А. Северцова24 «Динамика населения...» (1941) видно, что, например, низшие сумчатые вообще сумки не имели, а у высших она построена очень разнообразно. Плацента имеет у плацентарных очень разнообразное строение. Эти, видимо, общепризнанные факты позволяют уже утверждать, что плацентарные, сумчатые и т. д. характеризуются вовсе не плацентой, сумкой и т. д., а лишь способностью произвести сумку, плаценту и т. д. Причем эта сумка или плацента вовсе не предопределена как таковая, а лишь в форме потенции, могущей быть воплощенной самым разнообразным образом. Можно пойти еще дальше. Плацентой обладают не только млекопитающие, но и некоторые рептилии, а живородящие сцинки тупохвосты (Trachysaurus rugosus) обладают настоящей аллантоидной плацентой (Пузанов, 193825, с. 328, см. 121, 5*). Мне помнится, что плацента имеется и у некоторых акул, скатов и у некоторых живородящих насекомых. Любопытно было бы сравнить плаценты в столь широком сравнительно-анатомическом разрезе. С точки зрения кондициональ-
* Цифры 121.5 означают том 121 архива, позиция 5. (Примеч. ред.).
{55}
ных признаков весьма вероятно, что между некоторыми формами плаценты млекопитающих, акул, рептилий и пр. окажется больше сходства, чем между плацентами в пределах одного класса. В пользу того, что большинство признаков, характерных, например, для млекопитающих, окажутся кондициональными, говорит одно примечание в той же книге Северцова о динамике населения. Он пишет: или надо принять самостоятельное развитие важнейших признаков млекопитающих (волосы, млечные железы и т. п.) у разных групп млекопитающих, или допустить, что эти признаки были свойственны очень древней группе. Видимо, последняя альтернатива принимается эпигонами дарвинизма только в силу косности мышления.
Наконец, можно привести еще один прекрасный пример кондициональных признаков: расширение члеников передних лапок у самцов жуков. В самых разнообразных семействах (Carabidae, Tenebrionidae, Dytiscinae, Halticinae и т. д.) мы имеем роды, где самцы имеют расширенные лапки (причем с разным числом члеников), и другие роды, где расширения нет. Ясно, что это расширение произошло независимо в разных семействах и потому, может быть, для всех жуков кондициональным признаком является расширение члеников передних лапок.
Кондициональные признаки не могут рассматриваться как простое осуществление скрытого зачатка. Против этого говорит чрезвычайное многообразие проявлений кондиционального признака. Напротив, их скорее можно сравнить как бы с картиной, отброшенной проекционным фонарем на колеблющийся экран. Наилучшую аналогию этому можно найти в диалоге Платона «Государство» (в пещере на стене отражаются тени проходящих мимо людей). Идеи Платона могут иметь еще эвристическое значение и быть использованы для осмысления актов, примитивно толкуемых эпигонами дарвинизма (например, «девиации» А. Н. Северцова).
Понятие кондициональных признаков в значительной мере стирает границу между гомологичными и аналогичными органами. Я лично склонен думать, что понятие гомологии не носит обязательно генетического характера в смысле преемственности актуализированных форм, но может быть преемственностью в той потенциальной форме, динамическом эмбриональном поле А. Г. Гурвича26 и т. д., которую можно назвать гомологией высшего порядка. Само собой разумеется, что ведущие признаки нам придется отбирать из совокупности реальных, а не потенциальных признаков и этот путь отбора заключает в себе много исканий. Путь, который я проделал с личинками Melolontha и начал с видами рода Haltica, заключается в том, что для нескольких десятков признаков производится сравнение средних и их дисперсий. Здесь мы переходим к проблеме разграничения видов и других таксономических единиц.
2. Разграничение. Для простого описания ценность признака — в тесноте его корреляционных связей с другими признака-
{56}
ми (обилие коннотаций по Гельмтольцу). Для разграничения соседних таксономических единиц выдвигаются другие критерии. Здесь, как я уже указывал, всплывают вопросы о сравнительной ценности и иерархической подчиненности разных признаков. Эта сравнительная ценность может определяться разными критериями и вряд ли может быть выставлена a priori.
Чисто формальные критерии: первый — отсутствие или минимальная величина трансгрессии. В работе с Melolontha я применял очень простой критерий, названный мною коэффициентом дивергенции (в литературе этот термин применялся в другом смысле) и выражающийся формулой
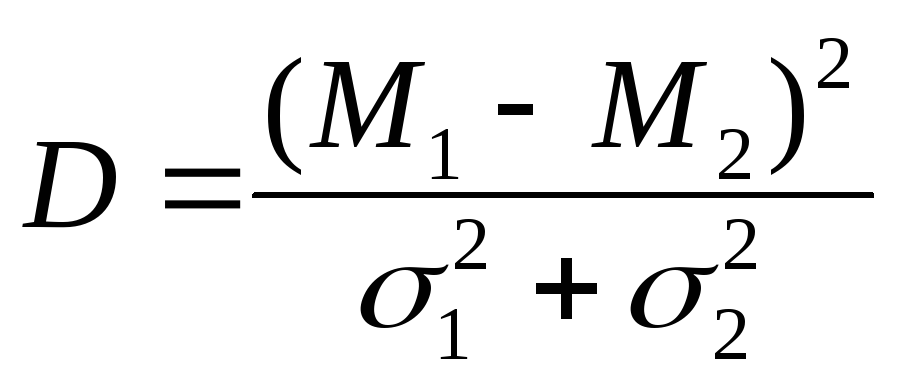
(квадрат разности медий, отнесенный к сумме дисперсий, квадратов сигм). Мы можем принять, что трансгрессия отсутствует (для простоты примем, что сигмы обоих видов равны), если расстояние между средними арифметическими равно шести сигмам (т. е. отклонение плюс три сигмы от одной средней как раз совпадает с отклонением минус три сигмы от другой). В таком случае этот коэффициент, очевидно, равен 18. В моей работе о личинках Melolontha «лучшие» признаки показывали коэффициент дивергенции, равный 0,8, и даже при таком малом коэффициенте комбинация нескольких признаков позволяла определять виды в большинстве случаев. Для ориентировочного определения реальности различий можно пользоваться таким упрощенным приемом: располагают исследуемый Признак в восходящий и нисходящий ряды обоих видов, затем считают число экземпляров обоих видов в области трансгрессии и относят его к общему числу экземпляров. Если процент (при общем количестве в несколько десятков) меньше 50, то можно уже с уверенностью говорить о реальности различия. Из взятых для испытания ряда признаков отбирают те, которые показывают наибольшие коэффициенты дивергенции. Эти уже отобранные признаки затем сводятся в комплексы по методам, намеченным Романовским и Фишером.
Второй формальный критерий — зависимость от экологических и географических различий. Эта зависимость имеет и недостатки, и достоинства. Признаки, зависящие от экологических и географических различий, конечно, мало пригодны для характеристики видов в целом, так как они увеличивают общевидовую дисперсию, но зато могут быть прекрасно использованы для определения географического положения и характеристики низших таксономических единиц.
Что же касается сравнительной значимости признаков, то по этому вопросу литература очень обширна, но обоснование тех или иных положений или очень туманно, или вовсе отсутствует.
Например, человекообразные обезьяны имеют те же группы крови, что и человек, и, видимо, можно переливать кровь от
{57}
обезьян человеку в пределах той же группы. По этому признаку того же «кровного» родства выйдет, что надо образовывать родственные группы, в каждую из которых войдут некоторые люди и некоторые обезьяны. Такую уродливую форму полифилетизма, я полагаю, никто защищать не станет. С другой стороны, можно привести ряд примеров, где правильная морфологическая группировка влечет за собой неизбежно и соответствующую биохимическую. Особенно показателен случай с комарами Culex и Anopheles. Различие родов было установлено, когда еще не была известна связь последнего рода с малярией, и, однако, изучение малярии вполне оправдало чисто морфологическую систематику. В противоположность биохимическим признакам, которым придается чрезмерно большое значение, есть признаки, которым отказывают в каком бы то ни было значении (например, окраска цветков по Линнею). Помню, А. П. Семенов-Тян-Шанский восставал против аналогичных утверждений энтомологов отказывать в таксономической ценности окраске. Он указывал, что есть такие группы, где окраска имеет определенное таксономическое значение. Даже у растений можно указать, например, на водоросли — бурые, зеленые, синезеленые, красные, где цвет является ведущим признаком крупных систематических единиц27. Но в общем Линней, конечно, прав: такие случаи, где цвет играет важную таксономическую роль — редкое исключение. На данном примере, пожалуй, мы действительно видим сохранение идей Линнея или Агассица о том, что определенным признакам свойствен определенный таксономический уровень. Копулятивные органы, по мнению многих, имеют особенно высокое систематическое значение, но неодинаковое таксономическое положение. Многие им приписывают особое значение для разграничения видов (в связи с общепринятым, хотя и не вполне правильным положением о бесплодии гибридов между видами), но не придают большого значения для различия высших систематических категорий (в ботанике имеет место другое, как уже было указано). Однако мы имеем и обратные случаи. Н. А. Теленга, например для браконид, выдвигал, что копулятивные органы очень важны для различения родов, но мало пригодны для различения видов. Для Halticinae, например, их пока применяли только для различения видов, и мой собственный опыт по ряду родов показывает, что при огромном разнообразии эдеагусов в пределах некоторых родов пока не представляется возможным выделить какие-то родовые признаки для копулятивного аппарата. Мало того, в строении эдеагусов мы наблюдаем конвергенцию не только в пределах рода (Longitarsus nigrofasciatus, L. brisuti), но и между родами: с эдеагусом этих двух видов очень сходен, отличаясь в общем только шириной, эдеагус довольно отдаленного рода — у Chaetocnema semicoerulea. Получающийся вывод о том, что конвергенция копулятивных органов имеет примерно то же распространение, что и в пределах других систем, надо тщательно проверить и сопоставить с высказыва-
{58}
ниями авторов, стремящихся доказать, с одной стороны, что по строению эдеагусов или внутренних мешков пенисов можно хорошо построить иерархическую естественную систему, являющуюся вместе с тем филогенетической, а с другой,— что имеет место тесная зависимость в развитии мужских и женских копулятивных органов (см., например: Даль — для пауков, Гольдхаус— для Microlestes из карабид, Жаннель — для Batiscinae28 и др. указания в одной из обзорных статей Гейкертингера в «Коleopterologische Rundschau» в конце двадцатых годов). Не исключена, таким образом, возможность, что, как правило, определенным ступеням системы будет соответствовать особая значимость различных признаков: окраска для некоторых категорий ниже вида, детали копулятивных органов для вида и т. д. Этим путем, может быть, удастся выделить истинное зерно в положении Агассица о таксономической неравноценности признаков.
Выявив сравнительную таксономическую ценность разных признаков на основании попыток «полного описания», можно будет приступить к решению задачи о дивергенции не только отдельных признаков, но и для генеральной совокупности признаков сравниваемых видов. Попытки в этом направлении уже делались, но как будто без достаточного продумывания самой возможности полного описания вида. Я помню статью Лахтина в «Антропологическом журнале» (в начале послереволюционных годов) о коэффициенте сходства. Особое внимание надо обратить на изучение пропорций тела и вообще на габитуальные признаки, находящиеся до сего времени в крайнем пренебрежении. Между тем несомненно, что они-то и руководят систематиками при разбиении на естественные роды и лишь потом подыскиваются морфологические признаки для большего удобства и для удовлетворения господствующей традиции. Конечно, вероятно, что роды отличаются друг от друга не только количественно, но и качественно в разных группах: например, весьма вероятно, что роды у ракообразных с морфологической точки зрения обширнее, чем у насекомых, у. млекопитающих — обширнее, чем у птиц. Роды у Halticinae в большинстве случаев очень естественны, характеризуясь и габитусом, и структурными признаками, но роды, группирующиеся около Podagrica или Crepidodera, часто характеризуются более формально: этот формальный подход, может быть, особенно резко выражен в подсемействе Galerucinae в группе родов, близких к Galeruca. Получив критерий сходства в общем виде, можно подойти к вопросу о количественном и качественном различиях родов в различных семействах и т. д.
3. Таксономические единицы разных порядков. По этому вопросу литература очень обширна, я же сам пока очень мало думал, так как, естественно, плодотворные мысли могут возникнуть лишь после значительной работы по оценке сравнительного объема и качественного различия разных систематических единиц. Для таксономических единиц ниже вида, конечно, надо проштудировать работу А. П. Семенова-Тян-Шанского, а для озна-
{59}
комления со взглядами генетиков — сводки Добржанского и Турессона29. Три единицы ниже вида довольно отчетливы: подвид (в основном определен географически), морфа (в основном определена экологически) и аберрация (определена исключительно генетически). Конечно, и подвид определен тоже генетически и при этом актуально, а морфа определена генетически кондиционально. (Здесь снова встречаемся с кондициональными признаками.) Очевидно, на очереди стоят вопросы о географической изменчивости видов, о сравнительном характере изменчивости разных таксономических единиц и т. д.
Синтетическая практическая (номографическая) систематика
Простейшее
применение мною номограмм к личинкам
Melolontha
показало, что они различаются (всегда
трансгрессивно) по ряду признаков, из
них два признака (число крючков
стридуляционного аппарата и число
преданальных шипиков) независимы один
от другого и от остальных трех (ширина
головной капсулы, длина 2-го и 4-го члеников
усиков), поэтому для них и для одного из
остальных трех (я брал ширину головной
капсулы — с)
можно применять метод Гейнке в его
первоначальном виде, остальные же
признаки надо брать с учетом их
коррелятивной зависимости. Положим
(рис. 1; линии регрессии, ограниченные
полосами ±3, показаны сплошными и
пунктирными линиями, а штриховкой
отмечена область трансгрессии видов А
и В), b
— длина 2-го членика усиков, c
— ширина головной капсулы. Тогда
![]() ,
где
— коэффициент регрессии, а буква с
черточкой наверху обозначает среднее
арифметическое данного признака. Квадрат
относительного отклонения наблюденного
значения от вычисленного будет равен
,
где
— коэффициент регрессии, а буква с
черточкой наверху обозначает среднее
арифметическое данного признака. Квадрат
относительного отклонения наблюденного
значения от вычисленного будет равен
![]()
где b — стандартное отклонение b от линии регрессии. Извлекая корень, получим
![]()
— линейная функция, для которой можно построить номограмму. Для экономии места я помещал шкалу с посередине, шкалы b и d (длина 4-го членика усика) — по бокам, а между шкалой с, принятой за независимое переменное, и шкалами b и d помещал шкалы обоих видов с отметками. Соединяя значения с и b для определенного экземпляра, читатель сразу прочитывал значение для данного экземпляра и прибавлял его к сумме по первым трем признакам. Подобным же образом (пользуясь правой частью номограммы; рис. 2) определялось значение и для d и с.
{60}
v
и по пяти признакам и дает материал для
суждения о принадлежности данной особи
к тому или иному виду. Чем больше эта
сумма, тем меньше вероятность принадлежности
данного экземпляра к данному виду. В
работе о Melolontha
я пытался определить рассеяние
эмпирически, но, мне думается, что если
v
получается от n
признаков, то
![]() для своего вида колеблется около единицы
со стандартным уклонением, равным
для своего вида колеблется около единицы
со стандартным уклонением, равным
![]() .
Это надо проверить на материале по
блошкам и посмотреть работы дельных
математиков, может быть Р. Фишера.
.
Это надо проверить на материале по
блошкам и посмотреть работы дельных
математиков, может быть Р. Фишера.
Конечно, такие номограммы могут быть построены и не для двух, а для нескольких видов. Кроме того, можно соединить в квадратной номограмме три независимых признака (рис. 3) и, пользуясь крестообразным транспаратном, прочитывать на четвертой стороне сразу сумму для трех признаков. Для оценки значимости рассеяния следует использовать функцию Эджвурта — Пирсона.
Я думаю развить метод номограмм в следующем направлении: положим, мы имеем четыре признака — а, b, с и d. Расположим значения их на четырех сторонах квадрата (рис. 4) и, соединяя попарно значения четырех признаков, по положению точки пересечения в поле того или иного вида судим о принадлежности данного экземпляра к тому или ино-
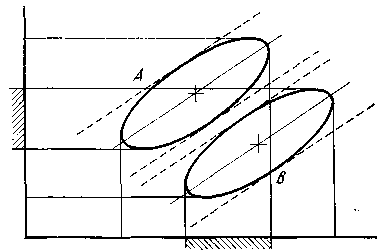
Рис. 1
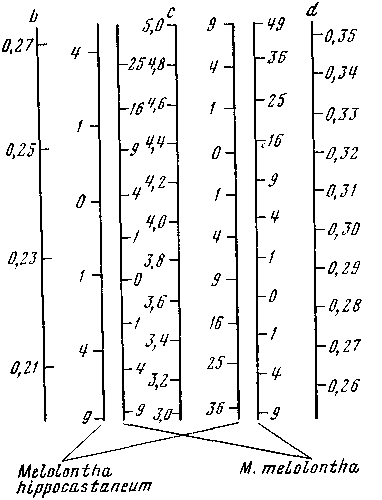
Рис. 2
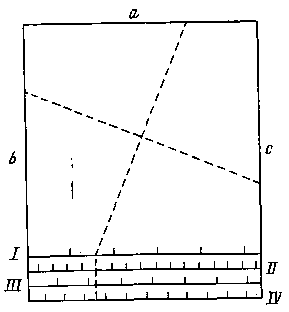
Рис. 3
{61}
му виду; на рис. 4 изображено поле для пяти видов. Концентрические эллипсы рассеяния соответствуют различным зонам значимости отклонения от центра вида. Степень надежности определения устанавливается по расстоянию точки пересечения от центра вида: чем меньше это расстояние, тем надежнее определение. Можно строить два квадрата для восьми признаков с целью контроля определения, а всего лучше построить гексагональную номограмму для трех пар признаков (рис. 5). В хорошо построенной номограмме три линии должны дать в пересечении маленький треугольник в поле одного из видов: если треугольник очень велик и если он не попадает в поле ни одного вида, то это значит, что или номограмма построена плохо, или мы имеем неизвестный вид. Из разговоров с М. Л. Франком, Л. Н. Бескиным и др. как будто выходит, что математики-номографы подобного рода номограмм не строили. Можно было бы помещать в каждую такую гексагональную номограмму лишь признаки одной плеяды (по терминологии П. В. Терентьева). Тогда три таких номограммы дали бы на трех страницах точное описание 18 признаков для целой серии видов: точность и сжатость описания чрезвычайно бы выиграли.
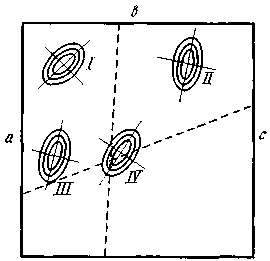
Рис. 4
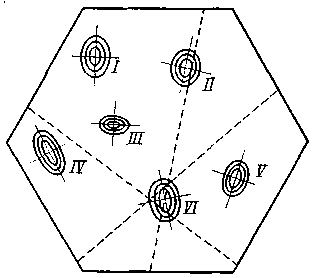
Рис. 5
ТЕОРЕТИЧЕСКАЯ ОБЩАЯ СИСТЕМАТИКА
Здесь я приведу только краткие общие соображения, поскольку эта часть должна развиваться в тесной связи с развитием практической общей систематики. Теоретическая общая систематика распадается по крайней мере на четыре ветви: 1) номотетическая систематика, 2) филогения и систематика, 3) логика систематики и 4) философия систематики.
{62}