

ipatov1997_fitotsenologija
.pdfТаблица 3.2
Пропускание ФАР одновидовыми синузиями нижних ярусов
елового леса, % от ФАР, поступающей к их пологу (по: Алексеев, 1975)
Вид |
|
Проективное |
покрытие |
|
||
|
|
|
|
|
|
|
|
40% |
60% |
|
80% |
|
100% |
|
|
|
|
|
|
|
Vaccinium myrtillus |
60 |
40 |
|
20 |
|
2-5 |
V.vitis-idaea |
45 |
25 |
|
12 |
|
4-6 |
Oxalis acetosclla |
65 |
47 |
|
27 |
|
10 |
Anemona nemorosa |
50 |
25 |
|
11 |
|
8 |
Fihpcndula ulrnaria |
35 |
15 |
|
8 |
|
1 |
Chamaenerion angustifolium |
45 |
25 |
|
10 |
|
1-5 |
Pieridium aquilinum |
— |
40 |
|
20 |
|
— |
Gymnocarpium dryopieris |
65 |
45 |
|
20 |
|
6-10 |
ATatteuccia, s truthiopteris |
— |
30 |
|
10 |
|
2-4 |
Equisetum. syhaticum |
50 |
30 |
|
18 |
|
10 |
Sphagnum girgensohnii |
— |
20 |
|
10 |
|
3 |
Polyirichuvi commune |
40 |
20 |
|
10 |
|
4 |
В пропускании света наблюдается определенная видовая специфика, обусловленная прежде всего различной площадью листовой поверхности. По данным разных авторов, листовая поверхность Pinus sytvestris в расчете на гектар составляет 1,2-5,7 га, Beiula pendula и B.pubescens 3,3-4,7 га, Picea abies 3,2-10,8 га. Строение крон, их ажурность у разных древесных пород неодинакова, поэтому при одной и той же сомкнутости пологами древостоев разных пород пропускается неодинаковое количество света (рис.3.5). Видовые различия отражены в ранжированном ряду некоторых древесных пород (ряд по уменьшению пропускания ФАР):
Рис.3.5. Пропускание ФАР (Т,%) пологом древостоев различного состава в зависимости от сомкнутости (С) при высоте солнца 35° ( по: Алек-
сеев, 1975).
1 — каменная береза, 2 - сосна обыкновенная, 3— березы бородавчатая и пушистая, 4 — ель европейская. 5 — бук восточный.
50

Береза каменная (Betula ermanii)
Сосна обыкновенная (Pinus sylvestris)
Береза бородавчатая (Betula pendula) и пушистая (В.pubescens)
Осина (Populus tremula)
Дуб летний (Quercus robur)
Лиственница даурская (Lartx dahurica) Ель европейская (Ргсеа abies)
Бук лесной (Fagus sylvatica) и восточный (F.orientalis).
Рис. 3.6. Пропускание ФАР по высоте.
А — буковый лес; 1 — под кронами, 2— в окне диаметром 2Ом. Б — сосняк грабово-буковый, сомкнутость 0.6-0.7; /— зона вершин, //— крон, 111— стволов, IV— бука и граба (по: Алексеев, 1973).
Рис. 3.7 . Пропускание ФАР по вертикали нижними ярусами ельника чернично-кисличного (по: Алексеев, 1973).
1 — с подлеском из рябины, ярусами кислицы и зеленых мхов; 2 с ярусом шиповника: 3 — с ярусом сфагновых мхов; 4 — с ярусом черники (пр.покрытие 80%); 5 — с ярусом кислицы (пр.покрытие 85%); 6— с ярусом зеленых мхов.
51

Рис. 3.8. Уменьшение освещенности по вертикали (по: Алексеенко, 1964; Kershow, Harris, 1971).
А — на лугах: I — с преобладанием клевера и тимофеевки; 2— овсяницы луговой; 3— ежи сборной; Б — в покрове из кустистых лишайников.
Распределение освещенности по высоте почти полностью определяется стратиграфией фотосинтезирующего аппарата. В буковом лесу (без подлеска и подроста с неразвитым травяным покровом) практически весь свет поглощается кронами бука на высоте 27-34м (рис.3.6). Основное поглощение света в грабово-буковом сосняке происходит в зоне крон соснового яруса и в ярусе, образованном буком и грабом. Аналогичным образом распределяется ФАР в нижних ярусах леса (рис.3.7). Характер распределения освещенности по вертикали на лугах, в моховом и лишайниковом покрове такой же, как и в лесу. Различается лишь высота растительного слоя, в котором происходит трансформация света: в лесу она измеряется метрами, на лугу сантиметрами, в лишайниковом и моховом покрове миллиметрами (рис.3.1, 3.7-3.9).
50 100
Освещенность,%
Рис. 3.9. Уменьшение освещенности по вертикали на лугах: (по: Кпарр,
1952; Donald, 1961):
1— с преобладанием крестовника, 2— с преобладанием трищетинника, 3— с преобладанием клевера, с плевелом (поле), 4 — с преобладанием белоуса и трищетинника.
52
Освещенность в толще растительного покрова изменяется в течение вегетационного периода в зависимости от развития фотосинтезирующего аппарата растений (рис.3.2, 3.10). В этом отношении резко различаются хвойные и лиственные леса; в первых световой режим почти не изменяется.
Рис. 3.10. Сезонные изменения пропускания ФАР (%) пологом южнотаежных древостоев при сплошной равномерной облачности (по: Алексеев,
1975).
1— березняк лещиново-грабовый, 2 — сосняк брусничный, 3 — ельник сфагново-черничный.
Очевидно, изменения светового режима растениями столь велики, что могут превышать пороги чувствительности практически любых видов. Отражение и поглощение основной части ФАР растениями верхних ярусов при их большой плотности приводит к тому, что нижерастущие получают скудный световой паек, и светолюбивые виды даже при прочих благоприятных условиях не могут здесь существовать. По данным В. А. Алексеева (1975), в ельнике-черничнике на долю ели приходится 83% общего количества ФАР, поглощаемого сообществом, а это практически вся ФАР, березы—7%, черники—3%, сфагнума — 5%, остальных — 2%;
в ельнике чернично-кисличном: ель поглощает 80%, береза—4%, |
ряби- |
||
на— 0,6%, |
щитовник австрийский (Dryoptens austriaca) — 0,3%, папоротник |
||
Линнея (Dryoptens linneana) — 1,0%, черника — 3,4%, |
кислица — 2,0%, |
сфаг- |
|
нум— 0,7%, |
зеленые мхи — 2%, остальные — 6%. |
Таким образом, древес- |
|
ный полог пропускает не более 10-15% ФАР, что и составляет световое довольствие растений нижних ярусов в этих лесах.
Большая плотность верхних ярусов в ельниках и буковых лесах препятствует развитию практически любых видов под их пологом, формируются так называемые мертвопокровники. Прекрасно развивающийся в разреженных сосняках вереск (Calluna vulgaris) из-за недостатка света не может расти при тех же почвенных условиях в высокополнотных сосняках. Мозаика светового режима, вызванная неравномерной плотностью растительного покрова верхних ярусов приводит к мозаике растительности нижерасположенных слоев. Исследования в Серебряноборском лесничестве (Малкина и др., 1970) показали, что самосев лиственных пород под материнским пологом в наиболее темных местах (пропускание света около 2%) погибает через 5-6 лет, в осветленных (при пропускании 7-10%) живет 14-15 лет. По сведениям Ю. А.Злобина (1960) при изменении освещенности в ельнике майниковом от 580 до 32700 люкс число подроста ели
53

увеличивается в 1,5 раза, а его проективное покрытие в 2,5 раза. Это приводит к неравномерности распределения древостоя, его мозаичности.
От светового режима, создаваемого доминирующими видами*, зависит видовое разнообразие сообществ. Например, в сходных экотопах в таволговых сообществах при полной сомкнутости таволги в три раза меньше видов, чем на лугу с преобладанием душистого колоска. В сообществах таволги число видов уменьшается с увеличением ее проективного покрытия. Ярким примером сезонных изменений нижних ярусов леса в связи с изменением светового режима служит развитие эфемероидов — пролески
(Scilla sibirica), хохлатки (Corydalis halleri), гусиного лука (Gagea lutea). Это происходит в весенний период в широколиственном лесу при обилии света, когда листвы на деревьях еще нет или она не полностью развернулась.
Следует иметь в виду, что одновременно с трансформацией света растения изменяют и другие экологические факторы, о чем будет сказано дальше, поэтому часто трудно выделить роль светового фактора во всем комплексе воздействующих на растения экологических факторов.
ТЕПЛОВОЙ РЕЖИМ
Тепловая энергия планеты почти полностью состоит из лучистой энергии солнца, незначительная ее доля приходится на тепловое излучение земли. Тепловая энергия, приходящая к конкретному участку растительного покрова, складывается из солнечной радиации, энергии, переносимой при перемещении воздуха, теплового излучения при окислительных процессах, в частности при разложении органики.
Рис.3.11. Зависимость величины альбедо (А) суммарной солнечной радиации некоторых типов фитоценозов от высоты Солнца, h (по: Руд-
нев, 1977).
1— заповедная некосимая степь, 2 — растительность большой лесной поляны, 3 — молодой смешанный лиственный лес, 4 — смешанный лиственный лес (50-60 лет).
Растительность трансформирует тепловую энергию, отражая, поглощая и выделяя ее. Отражение суммарной солнечной радиации растительным покровом незначительно (0,1-0,4%), зависит от высоты солнца,
*Доминирующий вид, доминант — преобладающий по обилию (проективному покрытию, числу особей, массе). Количественная граница между доминантой и недоминантом устанавливается обычно произвольно.
54

составам строения растительного покрова (рис.3.11). О тепловом режиме в растительном покрове можно судить на основании температуры тела растений, почвы, воздуха. С температурой связаны интенсивность физиологических и биохимических процессов, скорость биологического и химического разложения органического вещества, испарение и конденсация влаги и т. п.
Рис. 3.12. Суточный ход температуры воздуха в ясную (А) и пасмурную (Б) погоду (по: Чертовский, 1978):
1 — ельник-черничник; 2 — ельник травяно-сфагновый; 3 — открытое место.
Самая общая закономерность температурного режима в растительном покрове сводится к следующему. Удельная теплоемкость растительной массы выше, чем воздуха, поэтому поступающая в растительный покров энергия большей частью поглощается фитомассой, а на нагрев воздуха энергии расходуется меньше. В результате днем температура воздуха в растительном покрове оказывается ниже, чем на открытом месте. Ночью же при сокращении поступления солнечной энергии тепловая энергия излучается фитомассой и поглощается воздухом. В силу невысокой теплопроводности воздуха значительная часть тепловой энергии задерживается в растительном покрове и температура там оказывается выше, чем на открытом месте. В ельниках у поверхности напочвенного покрова в дневные часы температура на несколько градусов меньше, чем на открытом месте (рис.3.12). В пасмурную погоду днем эта разница меньше, чем в солнечную погоду, так как к растительному покрову поступает меньше тепловой энергии. В травяно-сфагновом ельнике температура воздуха ниже, чем в менее влажном черничном ельнике, видимо потому, что значительная часть тепловой энергии расходуется на испарение воды. Аналогично изменяет температуру травяная растительность. Так, если в 13 часов на лугу над поверхностью почвы при удалении травостоя температура оказалась равной 25,7°, то рядом на той же высоте под травостоем она была 19°.
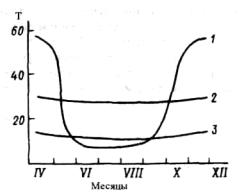
Во время вегетационного периода при вторжении холодных масс воздуха заморозки в лесу значительно слабее и менее продолжительны, чем на
55
открытом месте (рис.3.13). При слабых заморозках под растительностью у поверхности почвы обычно сохраняются положительные температуры, например на поверхности травостоя высотой 10 см наблюдались температуры -0,2, -3,2°, а на поверхности почвы соответственно +5,2 и +2,9°.
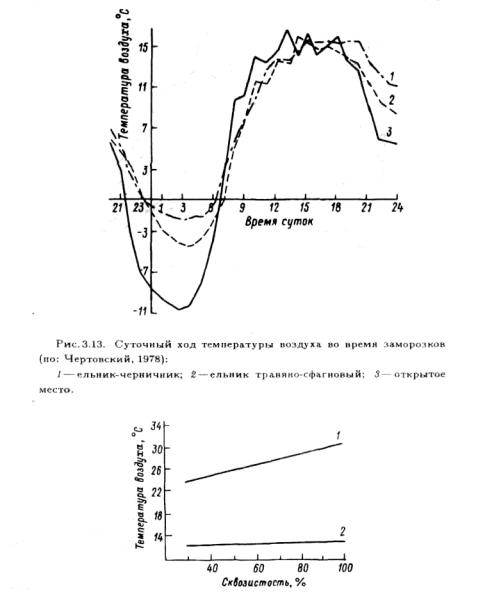
Рис. 3 .14. Зависимость температуры воздуха под пологом соснового древостоя от его сквозистости в ясную ( 1 ) и пасмурную (2) погоду.
Температура под пологом растений зависит от сомкнутости растений и фитомассы. Чем сомкнутость больше, тем ниже температура под пологом растений из-за того, что в этом случае в растительный покров проникает меньше лучистой энергии (рис.3.14). В пасмурную погоду, когда температура воздуха над растительным покровом снижается, разница температур над и под пологом оказывается незначительной.
56

Рис.3.15. Разность максимальных (А) и минимальных (Б) температур (ΔХ1) на поверхности мохового (1) и лишайникового (2) ковров и под ними при различной температуре воздуха (Т).
Таблица 3.3
Температура почвы на глубине 10 см на участках с оголенной почвой и люцерной (по: Шенников, 1941)
Воздушно-сухая |
|
Температура |
|
|
|
|
|
|
|
масса люцерны, |
максимальная |
|
минимальная |
средняя |
г/м2 |
|
|
|
суточная |
|
|
|
|
|
0(оголенная почва) |
30.4 |
|
18.0 |
24.2 |
90 |
26.5 |
|
18.4 |
22.4 |
310 |
18.6 |
|
16.5 |
17.6 |
При увеличении растительной массы на единицу поверхности увеличивается и поглощение тепловой энергии, в результате поверхность и верхние слои почвы нагреваются меньше, чем на оголенных участках (табл.3.3). При воздушно-сухой массе люцерны 3,1 т/га разница максимальных температур верхнего слоя почвы под люцерной и на открытом месте составила 12°. Существенную роль играет цвет растений. Поверхностный слой ковра зеленых мхов нагревается больше, чем светлых серовато-белых кустистых лишайников (рис.3.15). Вертикальное распределение температур зависит от расположения растительной массы. В дубовом лесу днем самая высокая температура воздуха наблюдается в зоне крон, несколько меньше и довольно стабильная под кроной, быстро уменьшается в слое подлеска и травяного покрова на высоте 1,5-0 м от поверхности почвы (рис.3.16). На лугу стратиграфия температурного градиента аналогична отмеченной в лесу (рис.3.17).
Растительный покров заметно меняет режим замерзания и оттаивания почвы по сравнению с оголенной поверхностью. Растительность, включая и образованный ею мертвый покров (подстилка, ветошь), играет роль теплоизолирующего слоя, поэтому под ней почва промерзает на меньшую глубину и оттаивает в весенний период медленнее, чем на открытых местах. Под мертвым покровом в заповедной степи глубина промерзания почвы в 1,5-2 раза меньше, чем на вспаханном поле (рис.3.18). А.А.Молчанов (1960) приводит многочисленные данные о промерзании
57

Рис. 3.16. Температура воздуха и почвы в дубовом лесу (изотермы в ясный день) (по: Горышина, 1969).
Н, см
Рис.3.17. Распределение температуры по высоте в заросли манжетки (день) (по: Л юбославский, 1916).
Штриховой линией отмечена высота кроны манжетки.
почв в разных лесах в сравнении с полем, покрытым снегом. Некоторые из них представлены в табл.3.4. В лесу промерзание почвы обычно меньше и сильно различается в разных типах леса. Так, в бору-долгомошнике оно незначительно в силу двух обстоятельств. Здесь имеется мощный теплоизолирующий слой из мха (Polytnchum commune), почвы сильно об-
58