

ipatov1997_fitotsenologija
.pdfформируются новые группы, с большими расстояниями и территориально больших размеров.
Имеет место еще одна тенденция. В результате более интенсивной дифференциации деревьев в группах и изреживания их с возрастом уменьшается неоднородность размещения, и распределение приближается к слу-
чайному (Бузыкин, 1967; Greig-Smith, Chadwick, 1965; Anderson и др., 1969).
Эта тенденция проявляется, в частности, в уменьшении варьирования наименьшего расстояния между деревьями с возрастом (табл.6.8). Если принять во внимание, что сухостой сохраняется на корню в течение 10-15 лет, то обнаруженное уменьшение коэффициентов вариации за это сравнительно короткое время можно считать значительным.
Таблица 6.8
Изменение наименьших расстояний |
|||||||
между деревьями с увеличением возраста |
|||||||
|
древостоя в сосняках |
|
|
|
|||
|
|
|
|
|
|
|
|
Древостой участков: состав. |
|
Класс |
|
|
|
|
|
бонитет, сомкнутость |
|
возраста |
n |
|
x |
u |
|
|
|
|
|
|
|
|
|
10С, II, 0.9 |
|
I |
252 |
|
39 |
52 |
|
|
|
|
271 |
|
57 |
53 |
|
10С, I, 0.8 |
|
II |
384 |
|
102 |
56 |
|
|
|
|
183 |
|
145 |
52 |
|
10С, I-II, 0.8 |
|
II |
500 |
|
64 |
68 |
|
|
|
|
150 |
|
134 |
5 5 |
|
10С, 1-11,0.6 |
|
II |
191 |
|
85 |
64 |
|
|
|
|
42 |
|
177 |
36 |
|
10С, I, 0.8 |
|
III |
476 |
|
68 |
65 |
|
|
|
|
244 |
|
94 |
58 |
|
10С, 11,0.4 |
|
IV |
264 |
|
183 |
50 |
|
|
|
|
167 |
|
205 |
45 |
|
10С, 111,0.8 |
|
III |
468 |
|
94 |
51 |
|
|
|
|
220 |
|
155 |
44 |
|
10С+Б+Ос, II |
|
V |
398 |
|
161 |
54 |
|
|
|
|
208 |
|
183 |
46 |
|
|
|
|
|
|
|
|
|
Примечание. Верхняя цифра — учтены все деревья, нижняя— только живые.
Таблица 6.9
Распределение Potentilla acaulis на 100 отрезках траисекты по пять площадок и сравнение его с биномиальным распределением (типчаково-тырсовал сухая степь, Семипалатинская обл.) (по: Василевич, 1969)
Число площадок с |
0 |
1 |
2 |
3 |
4 |
5 |
Potentilla acaulis |
|
|
|
|
|
|
|
|
|
|
|
|
|
Число отрезков трансекты |
23 |
26 |
22 |
12 |
10 |
7 |
Ожидаемые частоты |
10.6 |
29.9 |
34.0 |
19.3 |
5.5 |
0.6 |
Разница между найденными |
|
|
|
|
|
|
и ожидаемыми частотами |
+12.4 |
-3.9 |
-12.0 |
-7.3 |
+4.5 |
+6.4 |
|
|
|
|
|
|
|
199

Рис. 6.23. Распределение Festuca ovina no площади на разных стадиях сукцессии (по: Самойлов, 1984).
/— стадия 1 (гипотетическое распределение в прошлом); II: 1— стадия 2 (первоначальное распределение живых особей), 2 — стадия 3 (современное распределение); ///— стадия 4 (гипотетическое распределение в будущем). По оси абсцисс — классы расстояний, см; по оси ординат — частоты. Звездочками отмечены достоверные деформации кривых.
Выше приведены данные о распределении по территории (размещении) деревьев и отчасти кустарников. Иные жизненные формы -травы, мхи, лишайники подчиняются тем же законам. И у них обнаружены те же типы размещения. Регулярное распределение практически не встречается, преобладает контагиозное. Однако при изучении размещения травянистых видов возникают методические трудности, прежде всего с выбором счетной единицы. У многих видов без раскопки корневых систем не определить границы особей. Кроме того, при образовании парциальных кустов не известна степень их самостоятельности, иными словами, не ясно, насколько обеспечивается единое функционирование всего клона. В качестве счетной единицы можно принять надземный побег, дернину, клон, состоящий из парциальных кустов, или каждый куст в отдельности. При этом могут выявиться разные рисунки размещения. Таковы кривые распределения наименьших расстояний (рис.6.23) между особями овсяницы овечьей {Festuca ovina). У кривой П-2 статистически достоверно обнаружена деформация, свидетельствующая о том. что имеются два типа расстояний— в группах и межгрупповые. Кривая Н-1 деформации не имеет, размещение случайное. Кривая II-1 отражает результаты измерений между центрами клонов овсяницы: при построении же кривой П-2 учтены парциальные кусты. Достаточно эффективным и более простым оказывается использование территориальных единиц— мелких учетных площадок размером того же порядка, что и размер особей, либо несколько меньше (0,1, 0,5, 1м2). Учитывая присутствие-отсутствие вида на каждой площадке, построив на этой основе математическое распределение и сравнив его с соответствующим случайным распределением, можно сделать вывод о характере размещения изучаемого вида. Например, эмпирическое распределение Poientilla acaulis имеет повышенные частоты по сравнению с биномиальным в случае отсутствия вида (0) и его присутствия на четырех и пяти площадках в блоках по пять площадок (табл.6.9). Несомненно Potentilla acaulis распределена пятнами, имеет контагиозное раз-
200

Рис. 6.24. Трансекта в разнотравно-лабазниково-орляковом фитоценозе (Салаирский кряж, Новосибирская обл.).
1 — Pteridium aquilinum, 2— Filipendula ulmaria, 3 — Dactylis glomerate., 4 — Aegopodi-um podagraria, 5— Gtum urbanum, 6— Agrostis alba, 7— Pleurosfermum uralense, 8— Aconitum excelsiim, 9— Festuca pratensis, 10— Geraniujn pratense, 11 — Agrimonia pilosa. Стрелками показан возможный вариант разбиения трансекты на пятна доминирования (I-VI1).
мещение. В. И. Василевич (1969) приводит также сведения об аналогичном контагиозном распределении Cladonia rangiferina в сосняке лишайниковом с вереском и равномерном— Cytisus ruthenicus в сосняке сухотравнолишайниковом Казацкого мелкосопочника. Больше информации содержит учет проективного покрытия вида на мелких площадках (рис.б.24-6.26) (могут быть использованы и иные признаки — масса, число побегов). В частности, проективный учет позволяет выявить особенности варьирования вида в пределах пятен. Видно, что большинство видов размещено пятнами, кроме распределенных непрерывно по всей трансекте. Обнаружен
201

Рис.6.25. Изменение индекса различия и проективного покрытия некоторых видов (по трансекте) на пойменном лугу под одиночным дубом (р.Луга, Ленинградская область) (по: Прейс, Самойлов, 1977).
А — евклидово расстояние; Б — сглаженные проективные покрытия ви-
дов на площадках 0.1м2: 1— Trollius europaeus; 2— Geranium ■pratense; 3 — Galium boreak; 4 — Scorzonera humilis; 5— Phleum pratense; 6— Deschampsia caespitosa; 7— Alchemilla sp.\ 8— Centaurea jacea; В — то же, продолжение трансекты: 1 — Scorzonera humilis; 2— Convallaria majalis; 3— Leontodon his- ■pi&us; 4 — Alchemilla sp.; 5— Achyrophorus maculatus.
I-XI — порядковые номера контуров. По оси абсцисс — номера учетных площадок, по оси ординат — евклидово расстояние (А), проективное покрытие (Б, В), %.
202
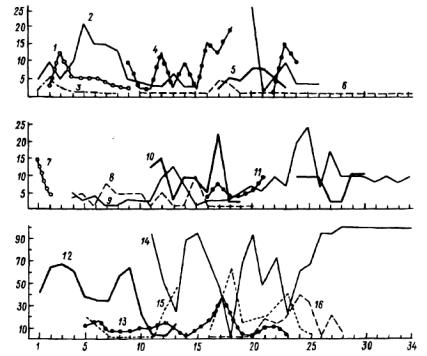
Рис. 6.26. Размещение видов (по трансекте) на окрайке верхового болота (центральная Карелия).
Виды: /— Vaccinium vitis-idaea, 2 — кустистые лишайники (Cladina arbuscula, C.rangiferina), 3— бокальчатые кладонии и политриховые мхи, 4.-— Betula папа, 5— Empetrum nigrum, 6— Oxycoccus ■palustris, 7— Arctosta-phylos uva-ursi, 8— Vaccinium. myrtillus, 9— Andromeda polifolia, 10— Chamae-dapkne calyculata, 11 — Ledum palustre, 12 — Calluna vulgaris, 13— Vaccinium uliginosum, Ц — сфагновые мхи (Sphagnum angustifolia, Sph.magellanicum, Sph.baHicum), 15— зеленые мхи (Pleurozium schreberi, Dicranum polysetum), 16— Eriopkorum polystachyon.
По оси абсцисс — номера площадок, по оси ординат — проективное покрытие, %.
высокий уровень случайного варьирования внутри пятен (рис.6.24). Вместе с тем обнаруживается и некоторая ущербность использования трансект из площадок, выстроенных только в один ряд. Если судить по рисунку, орляк (Pteridium aquilinum) размещен двумя пятнами. В действительности он представлен одним пятном, имеющим кольцевое строение. Трансекта пересекла в двух местах одно и то же пятно орляка и участок, лишенный его. Кроме пятнистости отражен клинальный характер (постепенное уменьшение) проективного покрытия по краям пятен (рис.6.25, 6.26). Особенно четко это заметно у сфагновых мхов и Calluna vulgaris (рис.6.26),
Trollius europaeus, Deschampsia cespitosa, AlchemiUa vulgaris (рис.б.25). Неко-
торые виды образуют пятна нескольких порядков (уровней): более крупные выделяются по присутствию вида, в пределах "пятен присутствия" заметны мелкие, различающиеся уровнем проективного покрытия. Крупные пятна Trollius europaeus и Alchemilla vulgaris состоят из более мелких, раз-
личающихся обилием этих видов (рис.6.25). Сфагновое пятно (рис.6.26)
203
можно рассматривать состоящим из двух пятен: на 28 -34 метрах трансекты сфагны имеют предельное значение обилия, и оно постоянно, в остальной части обилие колеблется случайным образом.
Причины возникновения пятнистости трав, мхов, лишайников, естественно, те же, что и деревьев: неоднородность экотопа и биотопа, неравномерность рассеяния зачатков, вегетативное размножение. Роль некоторых причин можно показать на приведенных выше примерах. Пятна сфагнума и шейхцерии обязаны своим появлением экотопу — они сформировались в зоне наибольшего обводнения. Пятна вереска, зеленых мхов, лишайников приурочены к песчаному склону, где сухо и бедно, а увлажнение происходит только за счет осадков. Вместе с тем зеленые мхи в этих условиях развились только благодаря притенению вереском — это причина биотопическая. Ю. И. Прейс и Ю. И. Самойлов (1977) убедительно показали, что основная причина мозаики в подкроновом пространстве дуба— неравномерная трансформация среды в разных его зонах.
Ранее, рассматривая вопрос о фитогенных полях растений, в сущности были раскрыты биотопические причины возникновения мозаики в размещении видов. Частичное наложение пятен орляка и таволги, видимо, объясняется тем, что почва одинаково благоприятна для них, а внутренний рисунок пятен других видов — в значительной мере совокупным влиянием на них орляка и таволги (притенение). В сухих сосняках на бедных песчаных почвах при восстановлений сосны зеленые мхи образуют пятна разных размеров. Можно отметить две причины пятнистости зеленомошного ковра: неравномерность притенения древесным пологом (в более притененных местах быстрее развиваются зеленые мхи) и разрастание мхов по периферии пятен, где они создают благоприятный для себя микроклимат.
Часто контагиозное распределение связывают с вегетативным размножением. Некоторые виды, размножающиеся семенами (например, Агпгса montana, Gentiana acaulis) распределены равномерно, а имеющие короткие корневища {Primula aunculata) — контагиозно. Ю. И. Прейс и Ю.И.Самойлов справедливо объясняют мозаику Scorzonera humilis, Convallaria majalis, Polygonum viviparum их клоновостью. Вместе с тем Ю. И.
Самойлов обнаружил, что на определенной стадии развития клоны Fesiuca ovina размещены равномерно. Таким образом, одной из предпосылок образования пятен является разрастание в результате вегетативного размножения.
Рисунок размещения видов не остается постоянным во времени. Типы размещения могут меняться в ходе сукцессии. X. Тагава (Tagawa, 1965) обнаружил, что контагиозное распределение может переходить в случайное и регулярное, а в ряде случаев вновь в контагиозное. Похожие изменения в стратегии овсяницы овечьей выявил Ю.И.Самойлов (см. рис.б.23). В заключение приведем данные Л. А. Маслова (1990). В разных типах леса практически все виды напочвенного покрова (злаки, разнотравье, кустарнички, мхи, лишайники) имеют разной степени выраженности контагиозное размещение. Всего им исследовано 62 вида, рассмотрено 143 случая и только в 14 из них обнаружилось в полной мере равномерное (случайное) распределение.
Неоднородность растительного покрова. Неравномерность распреде-
ления особей отдельных видов по территории отражает неоднородность растительного покрова как на уровне отдельных ярусов, так и всей его толщи. Если бы имело место независимое и случайное смешение разных популяций, то всегда наблюдалась бы очень большая пестрота растительного покрова. При этом число элементов мозаики, более или менее одно-
204

родных внутри себя, было бы весьма велико и превышало число видов на данной территории. В действительности имеются факторы, организующие смешение и распределение разновидовых комбинаций по территории, что приводит к упорядочению пестроты растительного покрова. Среди таких факторов — экотопические, фитогенные, зоогенные, естественно деструкционные, антропогенные.
Рис. 6.27. Микрокомплексность, связанная с микрорельефом и деятель-
ностью сусликов; Ергеня, верховья балки, Сталинградская область, 1950. (Съемка К.В.Зворыкиной; по: Лавренко, 1952).
Растительные группировки: 1 — Festuca sulcata-\-Stipa capillata (+Pyrethrum achilleifolium+Artemisia austnaca) в ложбинках 5-15 см глубиной, на светлокаштановой почве; 2— Festuca sulcata+Pyrethrum achtlleijolium+Stipa lessingiana (+Artemisia incana) на микроплакорах со светлокаштановой поч-
вой; 3— Роа bulbosa+Agropyrum ramosum (+Pyrethrum achilleifolium) в крайне незначительных понижениях, до 5см глубиной, на маломощной со-
лонцеватой светлокаштановой почве; 4 - Agropyrum desertorum^-Pyrethrum achilleifolium+Kochia prostrata (+Fcstuca sulcata) в очень слабых понижениях, 5- 10 см глубиной, или на едва заметных повышеньицах среди плоских пятен солонцов, на солонцеватой светлокаштановой почве; 5— Agropyrum desertorum+Kochia prostrata+Artemisia pauciflora (+ Artemisia incana) в слабо намеченных понижениях, на смытой солонцеватой светлокаштановой поч-
ве; 6— Kochia pro strata-\-Agropyrum desertorum (+Festuca sulcata+Artemisia pauciflora) в слабо намеченных понижениях, на корково-столбчатом солон-
це; 7— Artemisia pauciflora+Kochia prostrata в плоских понижениях, до 1015 см глубиной, на корково-столбчатом солонце; 8— Artemisia pauciflora+Camphorosma monspeliacum, в тех же условиях, что и предыдущая группировка; 9 — мелкие участки такыров, лишенные растительности; 10
— сусликовины с преобладанием Artemisia pauciflora; 11 — сусликовины с преобладанием Agropyrum desertorum и Festuca sulcata.
Если в пределах какой-либо территории экотопические условия различаются, то растительный покров на разных участках в результате экотопического отбора будет отличаться по составу и строению. Даже при выровненных условиях местообитания могут иметься небольшие микро-
205
повышения или западинки, площадь которых превышает иногда площадь питания крупных растений. В элементах такого микрорельефа в составе растительности встречаются растения, которых нет на основном фоне.
Конкуренция между растениями и трансформация ими среды обитания приводят к образованию ценоэлементов как в отдельных ярусах, так и во всей толще растительного покрова. Поскольку между соседними ценоэлементами имеются морфологические различия, они создают мозаичность растительного покрова. Мозаика растительного покрова возникает при интенсивном воздействии на среду животных (рыхление почвы и образование микрорельефа роющими животными, обогащение почвы под колониями птиц питательными веществами, неравномерное поедание растений и пр.). Иногда в результате деятельности животных создается очень пестрый растительный покров (рис.6.27).
Неоднородность растительного покрова создают и его естественные локальные разрушения. Например, ветровал, бурелом в лесу обычно приводят к формированию в так называемых окнах иной (отличной от основной) растительности, что связано с большим освещением в этих окнах. К мозаичности приводит и неравномерное выгорание нижних ярусов в лесу. Например, при беглом низовом пожаре пятнами разрастается вереск.
Человек проявляет себя по отношению к естественному растительному покрову обычно как деструктор. В лесу неоднородность растительного покрова возникает в результате вырубок деревьев, на лугах — неравномерного выкашивания травостоя. На месте старых стогов или кострищ формируется специфическая растительность.
Итак, неоднородность растительного покрова оценивается по внешним признакам, по его морфологии. Обычно элементы мозаики выделяются по обильным видам или группам видов.
Врастительном покрове (либо в отдельных его ярусах) можно выделять пятна доминирования. Границы такого морфоэлемента определяются по доминирующему виду или группе сходных видов, доминирующих в совокупности. В качестве сходных могут быть приняты растения разных видов, но одной жизненной формы. В сосновом лесу элементами мозаики, пятнами доминирования будут участки с господством зеленых мхов или кустистых лишайников. Можно выделить пятно доминирования, где диффузно распределены лисохвост луговой и ежа сборная. Основанием для их объединения служат кроме диффузного смешения сходство в размерах, положение фотосинтезирующего аппарата в одном слое, верхнее положение в травостое. Границу таких пятен проводят там, где изменяется обилие одного из видов или их совместное обилие.
Водно пятно доминирования могут быть объединены Scilla sibinca и Corydalis solida, поскольку они сходны по экологии. В пятно доминирования включаются и все остальные виды в его пределах.
Пятно доминирования может считаться однородным, но однородность эта относительна и условна. Дело в том, что пятно выделяется по обилию не всех, а части видов. Кроме того, при определении границы пятна мы должны принять условие — при каком изменении обилия можно считать, что пятно уже кончилось. Практически этот порог устанавливается не заранее, а в момент описания и может меняться в зависимости от конкретной ситуации. Более того, обилие видов в пятне доминирования оценивается
всреднем для всего пятна. Реально же обилие видов варьирует, и варьирование может быть особенно большим у видов, обилие которых не принимается во внимание при выделении пятна (рис.6.24). Два пятна (III и IV) могут рассматриваться не как однородные, а как клинальные. В
206
пятне III наблюдается клинальная комплементарность ежи и сныти, а в пятне IV — симметричная клинальность орляка и ежи.
Для выделения морфоэлементов, охватывающих все ярусы растительности (т.е. весь растительный покров), может быть использован тот же подход, что и при выделении пятен доминирования внутри ярусов. Однако необходимо принять во внимание "наложение" друг на друга пятен доминирования разных ярусов.
Назовем элементарную единицу неоднородности растительного покрова, охватывающую по вертикали весь покров снизу доверху, микрогруппировкой. Микрогруппировку можно определить как участок растительного покрова, внутри которого нельзя провести границы по заданным признакам и порогам, принятым для определения границы. При этом имеется
ввиду отсутствие границ во всех ярусах. Микрогруппировки могут быть относительно однородными и клинальными. Клинальная изменчивость может иметь место во всех (что бывает редко) или только в отдельных ярусах. Нетрудно заметить, что такое определение соответствует определению растительной группировки. Микрогруппировка и есть элементарная растительная группировка. Ее отличительная черта заключается
втом, что признаки и критерии ее выделения предельно (но разумно) детализированы. Границы устанавливаются по обилию видов во всех ярусах. Если в качестве основы выделения выбраны коллективные доминанты, виды объединяются по максимальному сходству. Так, участок леса, в одной части которого господствует осина, а в другой береза, может считаться растительной группировкой, если за основу выделения группировки принять господство мелколиственных пород в целом, а не отдельных видов. Но это не микрогруппировка. Участок же осинника с господством вей-
ника (CalamagrosUs arundtnacea) и костяники (Rubus saxatilis) на всей тер-
ритории, но с обилием орляка (Ptendium aquilinum) в одном месте и его отсутствием в другом представлен двумя микрогруппировками.
Примеры расчленения растительного покрова на микрогруппировки приведены на рис.б.27. Пятна доминирования, отраженные на рис.6.24, одновременно могут быть квалифицированы и как микрогруппировки.
Ценоэлемент по своему строению может соответствовать микрогруппировке. Но это бывает не всегда. Например, в дубняке можно выделить две микрогруппировки, различающиеся травяным покровом: в одной преобладает осока волосистая (Carex pilosa), в другой — обильны и осока, и сныть (Aegopodium podagrana). В микрогруппировки включен и древостой. Но поскольку различия в травяном покрове вряд ли связаны с эдификатором — ярусом дуба, то обе они входят в состав одного ультраценоэлемента. Аналогичная ситуация с латками крапивы и крупнотравья в сероольшанике. Выделение элементарных единиц в растительном покрове только на основе морфологии — упрощенный рабочий прием. Его можно использовать при массовых описаниях, при необходимости дать общую характеристику растительного покрова. В идеале членение следует проводить одновременно и по функциональным, и по морфологическим параметрам, тогда основными внутриценозными единицами придется признать ценоэлементы.
Фитоценоз
До сих пор говорилось о "внутреннем устройстве" растительного покрова на каком-либо участке. Естественно, возникает вопрос, а какова минимальная представительная единица растительного покрова? Обычно
207
элементарной территориальной единицей членения растительного покрова (растительности) принято считать фитоценоз.
Во введении мы уже определили фитоценоз как участок растительного покрова, устроенный по определенному закону, который может существовать самостоятельно вне данного окружения. В литературе накопилось большое число определений фитоценоза. Отвлекаясь от частностей, в большинстве из них можно выделить два компонента: указание на взаимодействия растений друг с другом и со средой и на однородность состава и строения. В историческом плане наибольшим признанием пользуются определения В. Н. Сукачева и А. П. Шенникова. Приводим эти определения. "Под фитоценозом (растительным сообществом) следует понимать всякую совокупность растений на данном участке территории, находящуюся в состоянии взаимозависимости и характеризующуюся как определенным составом и строением, так и определенными взаимоотношениями со средой" (Сукачев В. Н. Терминология основных понятий фитоценологии. Сов. ботаника, 1935, 5, с. 13). "Фитоценозом называется всякая конкретная группировка растений, на всем протяжении занимаемого ею пространства относительно однородная по внешности, флористическому составу, строению, по условиям существования и характеризующаяся относительно одинаковой системой взаимоотношений между растениями и со средой обитания" (Шенников А. П. Введение в геоботанику. 1964. с.12).
Эти два определения по существу не отличаются друг от друга, поскольку, исходя из иных определений В. Н. Сукачева и его разъяснений, можно утверждать, что под определенностью состава и строения он имеет в виду однородность. Разница сводится лишь к расстановке акцентов: в первом (определение В. Н. Сукачева) предпочтение отдается взаимодействиям, во втором (определение А. П. Шенникова) — однородности состава и строения. Наиболее обширным из дуалистических является определение В.Н.Сукачева, опубликованное в 1951 г. "1.Под фитоценозом или растительным сообществом понимается всякая конкретная растительность, на известном пространстве однородная по составу, синузиальной структуре, сложению и характеру взаимодействия между растениями и между ними и средой. 2. Фитоценоз охватывает всю растительность на этом пространстве, как высшую, так и низшую, включая и все микроорганизмы, относимые к растительному миру, т. е. фитоценоз включает всю — и надземную, и подземную растительность. Фитоценоз является сложным образованием, структурные части которого получили название синузий. 3. Фитоценоз представляет собой лабораторию, в которой происходит аккумуляция и превращение вещества и энергии. Взаимодействия между растениями, а также между всеми компонентами фитоценоза и между ними и средой связаны также с превращением вещества и энергии" (Сукачев В. Н. Основные принципы лесной типологии. Труды совещания по лесной типологии. 1951. С. 15). Во втором и третьем пунктах отражены два факта. Во-первых, фитоценоз, охватывая растения всей толщи растительного покрова, представляет собой не элемент, а собственно растительный покров. Во-вторых, как совокупность всех автотрофных организмов, занимая тем самым специфическое место в потоке вещества и энергии, он и с этой точки зрения является единицей растительного покрова. В нашем определении оба эти признака заключены в указании, что фитоценоз — участок растительного покрова, поскольку эти признаки — атрибуты растительного покрова.
Наличие взаимодействия между растениями и между ними и средой не специфично для фитоценоза, оно относится к широкому классу явлений,
208